










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Dental Journal
On-line version ISSN 0375-1562
Print version ISSN 0011-8516
S. Afr. dent. j. vol.76 n.6 Johannesburg Jul. 2021
http://dx.doi.org/10.17159/2519-0105/2021/v76no6a4
RESEARCH
The induction of bone formation: From bone morphogenetic proteins to the transforming growth factor-ß3 protein - Redundancy, pleiotropy and the induction of cementogenesis
U Ripamonti
MD, DDS, MDent, MFS, PhD (Med)., Bone Research Laboratory, School of Clinical Medicine - Internal Medicine, Faculty of Health Sciences, University of the Witwatersrand, Johannesburg, South Africa. ORCID Number: 0000-0002-6567-3594 Email: ugo.ripamonti@wits.ac.za
ABSTRACT
This review proposes to translate organogenesis and the induction of bone formation by the recombinant human transforming growth factor-ß3 (hTGF-ß3) in the Chacma baboon Papio ursinus to periodontal tissue induction and regeneration.
Naturally derived highly purified osteogenic proteins of the transforming growth factor-ß (TGF-ß) supergene family were implanted in Class II furcation defects of the first and second mandibular molars. Additional defects in P. ursinus were treated with recombinant human osteogenic protein-1 (hOP-1, also known as bone morphogenetic protein-7, hBMP-7) and hBMP-2, singly or in binary applications. In different studies defects were also implanted with hTGF-ß3 singly or in binary application with hOP-1. Harvested specimens on day 60 and 180 were processed for undecalcified histology using tungsten-carbide knives mounted on Polycut sledge' micro-tomes or the Exakt precision cutting and grinding system.
Highly purified osteogenic proteins showed the induction of Sharpey's fibres into newly formed cementoid with foci of mineralization. hOP-1 induced substantial cementogenesis whilst hBMP-2 preferentially induced alveolar bone. Intramuscular implantation of hTGF-ß3 absorbed onto coral-derived macroporous bioreactors engineered large heterotopic multicellular bone organoids.
Gene expression pathways by quantitative Reverse Transcription Polymerases Chain Reaction (qRT-PCR) show that the induction of bone is via several profiled BMPs and TGF-ßs expressed upon implantation of hTGF-ß3 recapitulating the synergistic induction of bone as shown by binary applications of low doses of hTGF-ß1 and hTGF-ß3 with hOP-1.
The rapid induction of bone by hTGF-ß3 provides the framework for a paradigmatic shift from recombinant hBMPs to hTGF-ß3 in clinical contexts, provocatively operational in periodontal tissue regeneration with substantial induction of cementogenesis in angiogenesis.
Keywords: Bone morphogenetic proteins' gene expression, qRT-PCR, TGF-ß3 master gene, noggin, molecular redundancy, pleiotropy, cementogenesis in angiogenesis, primates.
INTRODUCTION
Across the Century, systematic studies in the Chacma baboon Papio ursinus have reported the induction of cementogenesis with de novo nucleation of Sharpey's fibres inserted directly into mineralized dentine or newly forming cementoid matrix together with angiogenesis and capillary sprouting within the newly formed periodontal ligament space.1
Our first paper describing the induction of cementogenesis and of alveolar bone regeneration by highly purified naturally derived bone morphogenetic proteins (BMPs) in Class II furcation defects of Papio ursinus showed the substantial induction of periodontal tissue regeneration.2 We also showed that BMPs initiate cementogenesis, regulating the assembly of "a functionally orientated periodontal ligament" space.2 These preliminary observations in the non-human primate P. ursinus indicated that the presence of "multiple forms of BMPs may reflect a biological significance, locally regulating the regeneration of other tissues including the periodontal ligament and ce-mentum".2 This has suggested that the published material "may lay the foundations for systematic structure-activity relationships by recombinant hBMPs".2
These and other challenges were raised in a review that stated that the biological significance of redundancy of BMPs was a critical area of research ahead raising the question: "Does the presence of multiple forms of BMPs have a therapeutic significance?".3 It was also stated that the "future direction will depend on an optimal combination and/or developing a structure-activity profile amongst the members of the BMP family".2,3
The above challenges were addressed in a series of systematic experiments in P. ursinus aimed to reveal whether there is a structure-activity profile amongst BMPs family members, and whether the presence of multiple iso-forms may have a therapeutic significance.4-19
The aim of this review is to convey a perspective on the rapid induction of bone formation by the mammalian transforming growth factor-ß3 (TGF-ß3) isoform in Papio ursinus. As a prelude to the induction of periodontal tissue regeneration, the review begins by examining a number of classic experiments that, in our opinion, facilitated our current understanding of "Tissue induction"20 and of "Bone: formation by autoinduction".21 These seminal experiments helped to study the vast phenomena of tissue induction, morphogenesis, differentiation and de-differentiation, pleiotropy and redundancy. The multifaceted biological pathways highlighting the induction of bone formation are outlined below.
The review further translates the "operational reconsti tution" of the bone matrix22-24 and the "Bone induction principle"25 to periodontal tissue induction by the osteogenic proteins of the TGF-ß supergene family.26,27 Lastly, the manuscript reviews the molecular/morphological correlation of tissue induction and cementogenesis by the third mammalian TGF-ß isoform presenting novel data on vascular canals embedded into regenerated cementoid by the hTGF-ß isoform.
All the described experiments with the reported undecal-cified histology sections were performed in the Chacma baboon Papio ursinus. Animals were housed in the Wits Research Animal Facility (WRAF), Faculty of Health Sciences, University of the Witwatersrand, Johannesburg.
Selection criteria, housing conditions and diet were as described.2'4'9,28 For each different experiment, research protocols were approved by the Animal Ethics Screening Committee of the University. Experiments were conducted according to the Guidelines for the care and use of Experimental Animals prepared by the University complying with the Public Service Department National Code for Animal Use in Research, Education and Diagnosis in South Africa. Experiments were performed in consideration of the ARRIVE Guidelines.29
"Bone: formation by autoinduction"
"Bone: formation by autoinduction"21 epitomizes the development of regenerative medicine's dreams and paradigms. It was first invoked by postulating the presence of "mor-phogenetic factors", defined by Turing as "forms generating substances".30
Molecular and morphological observations have generated the high expectation of a regenerative scenario of "molecularly generated tissues and organs in assembling human tissue factories".31 Developmental molecular biologists, together with experimental surgeons and tissue biologists alike, are still dreaming of such "regenerative tissue and organ factories" to benefit the ageing human population, affected by debilitating disorders of tissue and organ failure together with all spectra of degenerative pathologies.31
Early research experiments pointed to the capacity of several extracellular matrices, including the uroepithelium (Fig. 1A),32,33 to initiate inductive phenomena, paradigma-tically set by the induction of bone formation.27 These experiments highlighted the induction of bone together with angiogenesis and capillary sprouting (Fig. 1B), to the extent that authors speculated the presence and release of putative morphogens, or morphogenetic signals, endowed with the striking capacity to initiate the heterotopic induction of bone formation.20-25
Trueta stated,34 "Angiogenesis is a prerequisite for os-teogenesis" and highlighted the "role of the vessels in osteogenesis" (Fig. 1B). In 1763, von Haller indicated that the "vascular system is responsible for osteogenesis", further stating that endothelial cells are osteoblasts' precur-sors.35 Much earlier, Aristotle associated sprouting blood vessels with a patterning function during organogene-sis.36,37 The patterning scenario envisioned by Aristotle's "morphogenetic vessels" highlights the vessels' capacity to act as a framework or "model" to shape the body structure (Fig. 1C). Recent work has expanded Aristotle's vision, providing evidence that endothelial cells are signalling centres secreting molecular signals in an angio-crine fashion to induce tissue morphogenesis.38
Senn 39 in 1889, used "decalcified bone, rendered not only completely aseptic, but thoroughly antiseptic by keeping it immersed for a considerable length of time in sublimate alcohol"39 to repair trephined defects in canine calvariae. Senn observed postnatal bone regeneration in treated defects upon implantation of decalcified bone matrices.
New bone developed via embryonic tissue induction, i.e. new bone formed recapitulating embryonic development.39 This recapitulation of embryonic development is pivotal to postnatal tissue induction, as we know it today: molecules exploited in embryonic development are re-exploited and re-deployed post-natally to initiate tissue induction and morphogenesis.26,27,40
In a number of reviews on the induction of bone formation,26-28, 45 we have always credited the experiments and conclusions of several authors, particularly highlighting the innovative work of Levander and his School in Köping.20,41,42 With a series of heterotopic experiments in lagomorphs, Levander shows an understanding of the basic mechanisms of the "bone induction principle",25 showing that, upon implantation of devitalized alcohol extracted bone matrices, bone forms directly from the mesenchymal tissue surrounding the graft.41,42
Levander states that "this specifically bone forming substance is liberated from the bone tissue and is carried by the tissue lymph to the surrounding areas where it is able to activate the mesenchymal tissue in such a way that this becomes differentiated into bone tissue, either directly or by means of the embryonic pre-existing stage of bone and cartilaginous tissue".41,42
In 1958, Moss described how an osteogenic inductor was extracted from bone.43 In his experiments, Gelfoam sponges incubated with bovine bone paste were implanted intracerebrally under parietal bone flaps in Long-Evans rats. Processed tissues showed extensive osteogenesis two weeks after implantation.43 Moss hypothesised that the intracerebral osteogenic activity was because of "stimulation of pre-existent osteoblasts in calvarial implantation sites by some factor extracted from the bone paste".43
The concept of "putative factors, substances and/or morphogens", firstly described by Turing.30 appears in the middle of last century epitomized by the innovative work of Levander and his team on alcohol-soluble osteogenic substance from bone.41,42 Indeed long before Urist' paper in Science,21 Levander postulated the presence of inductive substances within the solubilized matrix of bone.41,42 This was crystallized in his 1945 Nature' paper "Tissue induction".20
The hypothesis of putative substances and/or morphogens that initiate tissue induction is a fundamental step ahead from several experimentalists poised to Anally dissect the rules of tissue induction and morphogenesis. Amongst the firsts, Senn39 Sacerdotti and Frattin32 Huggins33 Levander,20,41 Moss,43 Urist,21 and Reddi,44 persevered to identify, purify and characterize the putative "osteogenic activity" present within several extracellular matrices yet endowed with the striking prerogative of de novo initiating "Bone: formation by autoinduction".21 The fascinating search for selected morphogens initiating the induction of bone formation allowed the discovery of an entirely new family of proteins initiators belonging to the transforming growth factor-ß (TGF-β) supergene family.26,27,40
Vessels and capillary sprouting are morphogenetic (Fig. 1C) but, after the fundamental studies of Trueta,34 are also "osteogenetic", i.e. capillaries that molecularly and morphologically regulate the induction of bone formation (Fig. 1B, C). The induction of bone formation is supported by an unlimited supply of responding perivascular pericytic and/or endothelial cells (Fig. 1B). Such "osteo-genetic vessels" express alkaline phosphatase activity and their basement membrane components bind angiogenic and morphogenetic proteins to provide the conceptual framework for the supramolecular assembly of the induction of bone formation defined as "osteogenesis in angio-genesis".26,27,45
Levander describes heterotopic bone induction as being rich in capillaries with multiple cells clustering around the invading vessels.20,41,42 In his experiments, Senn noted the importance of vascularised tissue in canine calvarial experiments and in clinical contexts.39 He described postnatal tissue induction as a recapitulation of embryonic development, preceding by almost 50 years the statements of Levander that "Post-natal tissue induction recapitulates embryonic development" and "the same substances deployed in embryonic development are re-deployed in post-natal tissue induction".20,41
Vukicevic et al. reported the in vitro development of long inter-connecting processes that resemble the osteocyte's canalicular network seen in bone when osteoblast-like cells were cultured on basement membrane components, namely type IV collagen, laminin, entactin and nidogen in a reconstituted Matrigel® Matrix.46 The authors further proposed that the osteocyte, a developmental stage of the osteoprogenitor-osteoblast lineage, retains a developmental 'memory' of the interaction between the osteoblast and components of the extracellular matrix of the invading capillaries, namely laminin and type IV collagen.46
This memory results in a ripple-like cascade of cell differentiation and the induction of bone formation.46 Reddi's incisive work46 has suggested that this "developmental memory" is re-activated by osteoprogenitors/osteoblastic cells reading laminin amino acid motifs across the basement' membrane components of the invading osteogene-tic vessels46 of Trueta' definition.34 The invading capillaries provide a framework for the newly forming bone made from mesenchymal condensations around each patterning capillary (Fig. 1C), and provide the foundation for the differentiation of osteoblastic cells.27,28,45
The multiple differentiating capacities of the endothelium, with its associated perivascular or pericytic cells, highlight the pleiotropic capacities of endothelial cells. A recent communication revitalizes the "osteogenetic vessels" of Trueta' definition,34 describing structurally distinct subsets of capillaries whose endothelial cells show specific expression profiles suggestive of specialised functional properties.47 The capillary endothelial subset, termed H endothelial cells, mediate localised growth of the vasculature providing niche signals for perivascular osteoprogenitors.47
How endothelial and osteoblastic cells communicate still remains little understood.47 The endothelial cell retains the plasticity of growth and differentiation with the capacity to de-differentiate into osteoprogenitors as well as a stem cell-like status for later rapid differentiation into both angiogenic and osteogenic phenotypes.38,47-49 Notch signaling promotes endothelial cell proliferation, establishing angiocrine signals, which couple angiogenesis to osteo-genesis with a further release of Noggin from endothelial cells, the former a selected antagonist of bone morpho-genetic proteins' signaling.50
Two experiments seemingly very different yet both highlighting the plasticity of the endothelium to differentiate and de-differentiate51,52 highlight such endothelial pleio-tropic plasticity. Aortic endothelial (E8) cells, treated in vitro with highly purified naturally-derived BMPs fractions51 showed prominent changes in the morphology of E8 cells (Figs. 1D, E, F). Identical concentration of BMPs fractions did not alter the cellular morphology of rat vascular smooth muscle cells (RVSMCs) over 24, 48 and 72 hours.51
E8 cells changed from a typical cobblestone appearance (Fig. 1D) to a spindle-shaped phenotype after 24 and 48 hours (Fig. 1E) with eventual rounding-up and detachment of cells by 72 hours (Figs. 1F). Reacquisition of the typical cobblestone appearance could be achieved by withdrawing BMPs fractions for more than 48 hours, provided the initial fractions did not exceed 6μg.51
The addition of highly purified osteogenic proteins fractions profoundly alters the endothelial phenotype (Fig. 1F), suggesting a critical role for bone matrix molecules in the phenotypic modulation of endothelial cells for the later induction of bone formation.51 Leversha et al.52 showed the capacity of glioblastoma (GBM) to generate stemness in tumour vasculature from putative cancer stem cells that comprise cell fractions capable of de-differentiation into endothelial progenitors.52 This may explain mechanistically why GBMs are amongst the most aggressive of human cancers, able to induce stemness in malignant cells that subsequently de-differentiate into endothelial cells52 providing nascent angiogenetic mechanisms for the survival and the rapid growth of the mother neoplasm.
The rapid induction of bone formation by htgf-ß3: morphological and molecular insights
Various delivery systems for soluble hTGF-ß3, applied singly or in binary synergistic application with osteogenic protein-1 (hOP-1/hBMP-7) were tested. These deliveries included allogeneic insoluble collagenous bone matrices (ICBM), calcium phosphate-based macroporous bioreac-tors, and Matrigel Matrix™.9,11,16,19,53-58
Histological analyses of hTGF-ß3 combined with insoluble collagenous bone matrix11 showed rapid induction of he-terotopically generated ossicles (Fig. 2A) with substantial corticalization of the newly formed mineralized bone (Figs. 2B, C). Newly formed cortices enveloped collagenous matrix remnants with limited, if any, vascular and cellular invasion, creating "tissue voids" within the central parts of the newly generated heterotopic constructs (Figs. 2B, C). Heterotopic ossicles are similar to the ossicles generated by synergistically combining hOP-1 with low doses of hTGF-ß154 or hTGF-ß3.16,53
With the evolution of more complex multicellular organisms, comes the need to establish an intricate system for cellular communication and control via signaling pathways such as the transforming growth factor-ß (TGF-β) supergene family.24,26,27,59 Insights into the molecular mechanisms underlying TGF-ß signaling have been garnered in organisms such as Drosophila melanogaster, Xenopus laevis60and Papio ursinus9-11,54-58In the latter experiments, we studied how the TGF-ß isoforms transduce signals in mammals and how they drive the induction of bone formation in primate species,16,54,55,58,61,63 including the sy-nergistic induction of bone formation when combining limited doses of either hTGF-ß1 or hTGF-ß3 with set doses of hOP-1/hBMP-7.16,53-58
Many TGF-ß activators have been identified.64-69 To transduce their signal, the TGF-ß ligands activate two classes of receptors64 such that the pleiotropic diversity in signaling by TGF-ß is achieved by various combination of ligand-receptor and receptor type coupling.64-69 Osteoblast differentiation by the TGF-ß superfamily is dependent on both the classical Smad pathway and Smad independent pathways.64-69 In Smad signaling, Smad complexes accumulate in the nucleus through the action of transcriptional regulators that direct coupling of the complexes to TGF-ß responsive promoters.67
A variety of transcriptional factor partners have been identified, and the transcriptional repertoire is finely tuned by the recruitment of co-activator or co-repressor proteins.67,70 Non-canonical Smad pathways include the ERK-MAP kinase (MAPK), JNK and p38 pathway.67,69,71 The p38 and Smad pathways converge to control RunX2 expression and osteoblast progenitor cell differentiation.70 TGF-ßs act as pro-migratory factors to mobilize and recruit stem cells from the surrounding tissue, which may also include peripheral blood.72 Multipotent mesenchymal stem cells (MSCs) are found in a variety of adult tissues, including muscle tissue and the periodontal ligament system.73,74 TGF-ß1 stimulates MSCs proliferation through the Smad3 pathway causing the accumulation of ß-catenin in the nucleus of responsive cells, thereby effecting gene expression profiles.75
Incisive studies have shown the peri-vascular origin of mesenchymal stem cells (MSCs),76 and reported that the MSCs found in several tissues and organs including the periodontal ligament system, are indeed pericytes.76 The authors concluded that the vascular walls harbor progenitor stem cells representing the peri-vascular or pericytic origin of all MSCs.76
Wang et al.77 showed that type IV collagens are important in establishing the Dpp dorsal-ventral gradient in Dro-sophila by supporting the assembly of Dpp complexes. This insight from Drosophila wing development patterning together with the observation of a marked increase in type IV collagen in generated ossicles by hTGF-ß3 in Papio ursinus61hints to the critical role that the extracellular matrix components play in establishing the pattern of activity required to induce bone by hTGF-ß3/treated macroporous bioreactors.
Until 1993, the prerogative of the induction of bone formation was assigned to the BMPs only.78 Research in the fruit fly Drosophila melanogaster showed high levels of homology between Decapentaplegic (dpp) and 60A genes with BMP-2 and BMP-4, and BMP-5 and BMP-6, respectively.27,78 This has suggested the primordial role of BMPs during the emergence and development of ver-tebrates.27,40
This common developmental role was shown by the capacity of recombinant dpp and 60A proteins to induce endochondral bone formation in heterotopic subcutaneous sites of the rat.79 Thus, a phylogenetically ancient signaling pathway deployed for dorso-ventral patterning in Drosophila, is also operational for the induction of bone formation in mammals.
The most appealing knowledge one can gather from the above findings is that Nature relies on common yet limited molecular mechanisms to provide the emergence of specialized tissue and organs, demonstrating evolutionary conservation of related proteins from phylogenetically distant species. Nature has usurped genes of the fruit fly operational for more than 800 million years to generate, with minor modifications of the amino acid sequence motif of the carboxy-terminal domain, the bone morpho genetic proteins, the induction of bone formation, skele-togenesis and with it, the emergence of the vertebrates and of the Homo clade.1
Both 125pg and 250pg hTGF-ß3 induce large heterotopic ossicles with bone initiating at the periphery of the calcium phosphate-based macroporous bioreactors (Figs. 2D, E, F; 3A; Figs. 4J, K).55,61-63 This occurs with the concomitant expression of members of the TGF-β su-perfamily, viz BMP-2, BMP-3, BMP-4, TGF-ß1, TGF-ß3and BMP-7, with however down-regulation of TGF-ß2 (Figs. 3A, B).55,61-63 A schematic of this proposed mechanism is shown in Figure 3.
The change in expression profile complements newly formed bone deposited by osteoblastic activity and osteoid synthesis (Fig. 3H). The newly formed bone shows prominent capillary sprouting and angiogenesis (Figs. 3H, I), the latter being marked by the increased expression of type IV collagen.61-63 Type IV collagen shows a significantly increased relative expression 15 days post implantation of hTGF-ß3 treated coral-derived macroporous bioreactors (Figs. 3A, B).55,61-63
At the molecular level, hTGF-ß3 treated macroporous bio-reactors induce bone by activating the BMPs pathway (Figs. 3A, B). Evidence of the BMPs expression pathway was shown in experiments where the inhibitory activity of the BMPs' antagonist Noggin was employed (Fig. 4).55,61-63 Hydroxyapatite coral derived macroporous bioreactors were loaded with binary applications of human recombinant Noggin (125 or 150 Mg).53,55,61-63 and 125pg hTGF-ß3, implanted into the rectus abdominis muscle of adult baboons and assayed for changes in gene expression patterns at 15, 60 and 90 days after implantation.55,61,62 The data correlated with bone formation by induction. Macropo-rous bioreactors treated with binary application of hTGF -ß3 and hNoggin had limited bone formation (Figs. 4C, D) with down-regulation of BMP2 and TGF-ß3 genes.55,61
The use of recombinant human Noggin (hNoggin) preloaded onto coral-derived macroporous constructs was also instrumental to mechanistically resolve the "spontaneous and/or intrinsic" induction of bone formation by coral-derived macroporous constructs when implanted in heterotopic sites of the rectus abdominis muscle of Papio ursinus.27,28,62Preloading macroporous constructs with 125 or 150pg hNoggin blocks the induction of bone differentiation.55,61 This has indicated that the spontaneous induction of bone formation by coral-derived macroporous constructs is via the BMPs pathway, since Noggin blocks the biological activity of the secreted and embedded proteins onto the substratum.55,61-63
Morphological analyses of tissues penetrating the macro-porous spaces of TGF-ß3-treated devices 15 days after implantation, show a microenvironment enriched with progenitor cells embedded into fibrin/fibronectin rings expanding into the macroporous spaces organizing tissue patterning and morphogenesis (Figs. 3C, D, F, G).55,61-63 The induction of fibrin-fibronectin rings is an early mor-phogenetic event signaling the differentiation of somatic invading cells into osteoblastic cell lines attached to and regulated by the developing extracellular matrix compo-nents.55,61,62 Extracellular matrix rings serve to structurally reorganize invading, migrating and differentiating mes-enchymal stem and somatic cells from the surrounding tissues to undergo differentiation and de-differentiation into osteoblasts,55,61,62 highlighting the importance of the "micro-environmental niche" in defining cell fate.53,58,63
Expanding extracellular matrix rings serve as anchorage for hyperchromatic cells interpreted as differentiating osteo-blasts re-programmed by hTGF-ß3 previously adsorbed onto the macroporous bioreactors from invading peri-cytic, perivascular myoblastic differentiated somatic cells (Fig. 3F blue arrows). RunX2 and Osteocalcin expression is significantly up regulated in hTGF-ß3 super-activated bioreactors on day 15 supporting the morphological observation of invading cells differentiating into the osteo-blastic phenotype with hyper-cellular osteoblastic activity and extracellular matrix secretion (Figs. 3H, I).
The importance of the microenvironment was highlighted by profiling the expression of TGF-β superfamily genes in the rectus abdominis striated muscle surrounding and enveloping the implanted macroporous bioreactors. Relative gene expression changes in TGF-ß1, -ß2 and -ß3 and BMP-2, BMP-3, BMP-4, BMP-6 and BMP-7 were assessed by quantitative Real Time PCR (qRT-PCR).
Sixty days after heterotopic implantation of hTGF-ß3-treat-ed bioreactors, there was increased relative expression of BMP-2, BMP-3 and BMP-4 with BMP-3 exhibiting the greatest change in expression, while BMP-6 expression was unchanged (Fig. 4B).55,61 Of interest, was the effect of the hTGF-ß3 treatment in the surrounding muscle tissue, which showed an increase in BMP-3 and BMP-7 although BMP-7 expression was decreased in the device when compared to untreated devices and control tissues.55,61
Data from these experiments also revealed the differential expression of the TGF-ß genes in response to implantation of hTGF-ß3 treated bioreactors. There was an increase in TGF-ß1 and -ß3 expression in both the device and muscle tissue relative to the control tissue and untreated control samples.53,61-63 TGF-ß2 expression decreased however in the treated devices but increased in the surrounding muscle tissue (Fig. 3B).
Morphologically there was extensive induction of bone formation by the 250pg hTGF-ß3 treated devices with bone forming at the periphery of the bioreactors only (Figs. 2D, E, F). Newly formed bone extendeding 3-4 cm from the profile of the implanted super activated macroporous bioreactors (Fig. 2F). Limited, if any, bone formed within the macroporous spaces of the implanted bioreactors (Figs. 2D,E,F).
The inductive capacities of the mammalian tgf-ß isoforms in preclinical contexts
Morphological and molecular data have shown that in different animal models including rodents, lagomorphs, canines and pigs, heterotopic implantation of the three mammalian TGF-β isoforms does not result in bone differentiation by induction.11,26,27,79 Klar et al.55 suggested that the "molecular redundancy" associated with "different molecular functions in primate tissues represents fine tuning of speciation-related molecular evolution in anthropoid apes at the early Pliocene boundary which resulted in a more capable tuning of the bone induction cascade in primate species".55
Following identification of the first TGF-ß1isoform in human platelets,80 several reports proposed, but did not show, that the transforming growth factor-β proteins might possess inductive activities as well as bone forming capa-cities.81-84 In an in vivo model in neonatal rats, injection of either hTGF-ß1or -β2directly into the periostea of the parietal bones resulted in a 2-fold increase in bone thickness in a dose-dependent manner.83 Further studies by Sporn and colleagues proposed that the TGF-β was an "initiator" of chondrogenesis and osteogenesis in the rat femur.84
Initiators, or inducers, of bone formation are different from factors that may promote and/or maintain tissue induction cascades. An osteoinductive protein must be capable "of initiating endochondral bone formation in heterotopic ex-traskeletal sites of animal models".11,21,22 Heterotopic sites avoid "the ambiguities of the orthotopic site where some degree of bone formation by conduction may occur from the viable bone interfaces".21 Direct injection of morphogens isolated from naturally derived bone matrix or platelet extracts, interact with developed cellular populations and differentiating progenitors in the subperiosteal regions of either calvariae or long bones that are amenable to the rapid promotion of bone formation.83,84
The molecular cloning of the osteogenic proteins of the TGF-β supergene family26 and the results of numerous preclinical studies in mammals including non-human primates have prematurely convinced molecular biologists, tissue engineers and skeletal reconstructionists alike to believe that a single recombinant hBMP would result in clinically acceptable tissue induction and morphogenesis in human patients. This theoretical potential has not been translated to acceptable results in clinical contexts.
Clinical trials in craniofacial and orthopedic applications such as mandibular reconstructions, sinus-lift operations and tibial non-union have indicated that supra-physiological doses of a single recombinant human protein are needed to induce often-unacceptable tissue regeneration whilst incurring significant costs without achieving equivalence to autogenous bone grafts.27,28,45,62
A recent study Anally reported that supra-physiological doses of hBMP-2 were required to induce often clinically insufficient bone in clinical contexts, and recognized potential local and systemic adverse effects.85 A proposed BMP/activin A chimera was formulated with superior activity to native BMPs, i.e. protein fractions extracted and purified from animal bone matrices,85 in inducing bone in non-human primates with reduced doses compared to the BMP-2/absorbable collagen sponge previously approved for clinical use.85 The recombinant chimera needs now to pass through clinical trials prior to seeking FDA approval.
A series of systematic studies in the non-human primate Papio ursinus showed that the hTGF-ß3 isoform is the most powerful soluble osteogenic molecular signal of the TGF-β supergene family so far tested in primates.11,55,58,61,63 The robust induction of bone formation by hTGF-ß3 when pre-combined with inactive insoluble bone matrix11 or coral-derived macroporous bioreactors55,61 has required the re-evaluation of the induction of bone formation in primate models.18,62
The use of insoluble collagenous bone matrix as carrier results in the rapid induction of bone formation (Fig. 2A) with corticalization of newly formed ossicles as early as 30 and 90 days after heterotopic implantation with explosive growth at the periphery of the implanted specimens (Figs. 2B,C).11 This explosive peripheral pattern is also seen when hTGF-ß3 is combined with macroporous calcium phosphate-based bioreactors (Figs. 2D, E), transfiguring the surrounding rectus abdominis muscle (Fig. 2F).58
The biological rationale for this explosive growth, supported by qRT-PCR data, is that the super-activated bio-reactors rapidly transform progenitor cells of the rectus abdominis muscle via gradient activities enveloping the implanted bioreactors as initiated by the recombinant mor-phogen (Figs. 2D, E, F, 3A, 4J, K).53-58
Additional systematic experiments in the rectus abdomi-nis heterotopic intramuscular sites of Papio ursinus have shown that the induction of bone by hTGF-ß3 applied singly is equal if not greater when compared to hetero topic ossicles generated by the synergistic induction of bone formation with binary applications of hOP-1/hBMP-7 with relatively low doses of hTGF-ß1 and -β3.16,53,54
Because of the pleiotropic biological activity of the recombinant hTGF-ß3 in different orthotopic and heterotopic sites for craniofacial reconstruction,58 experimentation to transfigure neoplastic carcinomatous masses was deemed a necessary experiment. This also followed the evidence of tissue transfiguration of the striated rectus abdominis muscle after heterotopic intramuscular implantation of 250 hTGF-ß3 combined with either collagenous matrices or coral-derived macroporous bioreactor.58,62
Bioptic material from harvested human squamous cell carcinomas (hSCCs) were transported in sterile medium to the laminar flow unit of the Central Animal Services (CAS) of the University Faculty of Health Sciences. Human and animal ethics clearances were obtained from the University (Human Research Ethics Committee Clearance certificate No. M150608). Fragments of 3 to 5mm in size were transplanted bilaterally under skin of the chest; bilateral pouches were created with sharp and blunt instruments to accommodate the transplanted hSCCs human biopsies' fragments.86
hSCCs grew within to three weeks to reasonable sizes. Tumors growth was palpated and when to sizeable sizes, nude mice under anesthesia were palpated in the thoracic area and approximately 300 to 500pl of Matrigel®Matrix with 250pg hTGF-ß3 were injected into the right neoplastic masses. In a number of mice, masses sustained multiple injections up to three injections of Matrigel®Matrix with 250pg hTGF-ß3 before euthanasia and tissue harvest.
Histological examination of resin-embedded sections of hTGF-ß3 injected hSCCs transplanted under the skin of the chest, showed a reproducible recurrent histological pattern of undifferentiated anaplastic growth at the periphery of the transplanted SCC biopsies vs. a different yet reproducible pattern of a highly differentiated oncotype in the center of the transplanted and harvested specimens.86
This oncotype and pattern' variations are of great significance. The morphological data show reproducible patterns of growth spatio/temporally distributed, i.e. poorly differentiated up to anaplastic SCCs at the periphery of the transplanted biopsies vs. more differentiated with keratinized oncotype in the center of the injected growing carcinomas, thus less malignant and more differentiated oncotype in the center following injections of doses of hTGF-ß3 in the center of the neoplastic masses using the Matrigel®Matrix delivery system.86
Histological examination of transplanted and injected SCCs showed that hTGF-ß3 injections into hSCCs induced an oncotype characterized by a shift into highly differentiated oncotypes with multiple pearls of keratinization, with less neoplastic undifferentiated cells. Injected hTGF-ß3 vs. non-injected hSCCs showed a remarkable reproducible onco-type variation from poorly anaplastic undifferentia-ted to highly differentiated and keratinized oncotypes after hTGF-ß3 injections in Matrigel®Matrix.86
As a final mechanistic note, the powerful transfiguration patterns as seen within the center of the hTGF-ß3 injected hSCCs heterotopically growing and expanding subcutaneously in athymic nude mice show that the injected soluble molecular signal in Matrigel®Matrix activates the cellular memory of the transplanted carcinomas. Cancers survive by recapitulating mechanisms of the normal development. The injections of relatively high doses of hTGF-ß3 in Matrigel®Matrix re-introduced a memory of developmental events already known to the affected cells, bringing back affected cells to their initial nonneoplastic and keratinized status.86 The transfiguration mech-anism(s) by hTGF-ß3 in Matrigel®Matrix set into motion gene expression pathways bringing neoplastic cells back to their initial stage with keratinized pearls of a highly differentiated oncotype. Collectively, that available data show that hTGF-ß3 locally initiate tissue transfiguration in vivo yielding highly differentiated oncotypes.86
In studies using isolated transitional epithelial fetal baboon bladder cells cultured on Matrigel Matrix™, we reported the generation of three-dimensional constructs of transitional epithelial cells with branching morphogenesis and lobulation of replicating transitional epithelial cells, which we defined as spheroidal organoids.87 Organoids are three-dimensional constructs composed of multiple cell types that originate from stem cells by mean of self-organization capable of simulating the architecture and functionality of native organs.88
The induction of a three-dimensional in vivo culture system combining the morphogenetic soluble signal of the recombinant hTGF-ß3 with a coral-derived macroporous bioreactor, morphogenizes the rapid induction of de novo bone organoids within the rectus abdominis muscle of Papio ursinus.
These in vivo multicellular organoids of mineralized bone, with osteoid and rapidly expanding marrow cavities, show regional anatomical and molecular identities organized as discrete domains temporo-spatially separated by morpho-genetic gradients.58,61-63 Organoids form by recruiting peripheral stem and/or somatic cells and de-differentiating myoblastic, pericytic and perivascular somatic cells into newly differentiated osteoblasts that induce rapid bone formation.61-63 Molecular gene expression analyses directly correlate with the anatomical morphological domains, showing a ripple-like cascade of changes in gene expression at the periphery of the intramuscularly implanted bio-reactors (Fig. 3).61-63 The implanted bioreactors per se, particularly at early time points, show limited cellular invasion within the macroporous spaces with minimal, if any, up regulation of morphogenetic genes.58,61-63
Whilst the paper of Alliston and Derynk89 misses to report the evidence of the induction of bone formation by the mammalian TGF-β isoforms,11,26,54,90 it shows that TGF-β "is a key regulator of the mechanical properties and composition of the bone matrix".89 Our studies in the Chacmababoon Papio ursinus show the spatio/tem-poral distribution of gene expression pathways across boundaries of the in vivo generated organoids. Anatomical morphological boundaries are the muscular pouches and the linea alba of the rectus abdominis muscle, and particularly so, the macroporous configuration of the super activated bioreactors set by the exoskeleton of the coral-derived constructs. Our systematic studies have shown how TGF-ß3 controls the developmental cascade of the induction of bone formation in primates.53-58,61-63
The research work from laboratory' benches to pre-clini-cal studies in Papio ursinus has been translated to a long term follow-up of pediatric patients following reconstruction of massive mandibular defects with recombinant hTGF-ß3.91 Ongoing experiments profiling additional pathways acting in concert with the master regulator TGF-ß3 will yield important information on the complexities underlying the induction of bone formation by hTGF-ß3 in the Chacma baboon Papio ursinus with relevance to man.
How can this wealth of knowledge be applied to perio-dontal tissue regeneration, and to the induction of cemen-togenesis with de novo insertion of Sharpey's fibers, the essential ingredients to engineer new attachment formation?
Cementogenesis by htgf-ß3and homologous morphogens of the superfamily
The operational reconstitution of a soluble signal with an insoluble signal or substratum 22,23,27,28,40 enabled the identification of soluble osteogenic molecular signals within the bone matrix. It has resolved the biological background of the osteogenetic capacity of the bone matrix.22-24
The dissociative extraction and reconstitution of a soluble signal with an insoluble signal or substratum restored the biological activity of the chaotropically extracted de-mineralized bone matrix.22-23 This operational reconstitution additionally defined the heterotopic bioassay,22,23,26-28,40 and the bases for translational research of the "Bone induction principle"25 to pre-clinical and clinical contexts.27,28,40,45,91
How the foreground review on the molecular and biological activity of the hTGF-ß3 isoform can be applied, modified and tuned to achieve the induction of cementogenesis, angiogenesis and capillary sprouting with de novo induction of Sharpey's fibers inserting into newly formed as yet to be mineralized cementum or cementoid?
The rational for linking "Bone: formation by autoinduction"21 to periodontal tissue induction is based on the discovery of the pleiotropic activity of the osteogenic proteins of the TGF-ß supergene family.9,26,27 Such proteins "provides soluble osteogenic molecular signals endowed with the striking prerogative of initiating cementogenesis and the assembly of newly formed ligament fibers in primates".1-3,9,14,15
We further quote verbatim a critical statement of the induction of bone formation and regenerative medicine at large that has propelled forward the "Bone induction prin-ciple"25 from preclinical to clinical contexts: "Obedient to the classic evo-devo' rule, developmental events that initiate in embryogenesis can be redeployed and recapitulated post-natally in tissue induction and thus rege-neration".1
Several studies have shown that the embryological phases of craniofacial, periodontal and tooth morphogenesis display multiple mRNAs as well as secreted morphogenetic and osteogenetic proteins species predominantly of the TGF-ß supergene family.1,7,9 Such genes and secreted gene products, members of the TGF-ß supergene family, singly, synergistically and synchronously initiate tissue induction and morphogenesis of the periodontal tissues and other craniofacial structures (Fig. 5).1,7,9 The immunolocalization of distinct morphogenetic proteins within the periodontal ligament space of 12- to 18-day-old mouse heads showed that there is a site specific localization of immu-nolocalized BMPs molecularly highlighting the biological significance of redundancy as well as the structure/activity profile of each tested protein (Figs. 5, 6).1,7,9,18
Developmentally, in situ hybridization of murine craniofacial structures, including tooth morphogenesis, with osteogenic protein-1 (OP-1, BMP-7) shows expression of OP-1 in the dental epithelium and the dental mesenchyme of developing murine molars (Fig. 5A).92,93 In context of the perio-dontal ligament space including cementum and alveolar bone, BMP-2 is primarily osteogenic with limited if anycementogenic inductive activity.7-9Assuch, immunolocalization of BMP-2 is strictly confined to the alveolar bone (Fig. 5B) with lack of immunolocalization within the ce-mentum and the periodontal ligament space (Fig. 5B). The immunolocalization pattern of BMP-2 reflects the structure/activity profile of the isoform with the substantial induction of alveolar bone both in canine and non-human primates' models with limited if any cementogenic capacity.7-9
To the contrary, osteogenic protein-1 (OP-1), when applied to periodontal defects created in the Chacma baboon Papio ursinus, is preferentially cementogenic, with the induction of Sharpey's fibers within newly formed cementum along the planed root surfaces.1,7-9,15,18 Once again the immunolocalization pattern of the OP-1 isoform defines the structure/activity profile of the protein (Figs. 5C, D, E). OP-1 was found to be expressed in newly secreted ce-mentum and cementoblasts in 16-days-old mouse pups (Fig. 5C) with a strong signal in predentine and mantle dentine (Fig. 5D).7
OP-1 Immunolocalization of the periodontal ligaments space on 18-days-old mouse pups shows pronounced selected immunostaining of the newly formed cementum with a trabecular pattern of expression across the perio-dontal ligament fibers with minimal if any immunolocali-zation to the alveolar bone (Fig. 5E). The composite expression of molecularly different yet homologous morpho-genetic proteins has suggested that optimal therapeutic regeneration may entail the combined use of homologous yet molecularly different proteins.1,7-9,15,18
Particularly enthralling are the expression patterns of BMP -3 gene and gene product in homogenized generated ossicles by the heterotopically implanted coral-derived macroporous bioreactors.55,61-63 The pleiotropic activity of the BMP-3 gene and gene product is shown in Figure 6. The protein is immunolocalized throughout the periodon-tal ligament space including the alveolar bone and the cementum (Fig. 6A).7 Of note, BMP-3 immunolocalizes along the inferior alveolar nerve (Fig. 6A white arrow).7
Surprisingly, as it may seems for a bone morphogenetic protein,27,28 BMP-3 immunolocalizes in the cerebellar white matter of the cerebellar folia (Fig. 6B).7 BMP-3 immunolo-calizes in the cytoplasm of Purkinjie cells of 13-days-old mouse pup (Fig. 6B)7 delineating the axonal neurite of the Purkinjie cells (Fig. 6B).7
These findings by Thomadakis et al.7 are noteworthy, and indicate that BMP-3 is neurotrophic during development and maintenance of the mammalian nervous system. Potentially thus, BMP-3 immunolocalization in the cerebellar folia and in the cytoplasm of Purkinjie cells and neurite axons may simply control the cerebellar fine tuning of striated muscle activity (Bone Research Unit, unpublished data 1998).
Immunolocalization of BMP-3 in the periodontal ligament space, alveolar bone and cementum (Fig. 6A) has indicated that the isoform may recapitulate post-natally the induction of all the essential components of the peri-odontal ligament system, i.e., cementum, Sharpey's fibers, periodontal ligament fibers and the alveolar bone. This hypothesis was tested by implanting highly purified osteo-genic proteins fractions, predominantly containing BMP-3 as shown by partial amino acid sequence information2,27 into Class II furcation defects of adult Chacma baboons Papio ursinus.2
Defects were implanted with 150 mg of baboon insoluble collagenous matrix (ICBM) reconstituted with 250μg of highly purified osteogenic fractions after gel filtration chromatography on Sephacryl S-200. Fractions were purified greater than 50.00-fold with respect to initial crude extract.2 This naturally derived highly purified preparation is predominantly composed of BMP-3 (osteo-genin)2 and BMP-2, with no detectable TGF-ß1 or TGF-ß2 (NS Cunningham and AH Reddi, unpublished data).
Implanted defects were harvested on day 60 after implantation as undecalcified blocks containing the first and second mandibular molars surrounded by periodontal tissues.2 Specimen blocks were embedded undecalcified in methyl methacrylate (K-PLast, Medim Germany).
Undecalcified serial sections including dentine and associated periodontal tissues were cut at 3 to 6 μm in the mesio-distal plane throughout the entire bucco-lingual extension of the furcation defects using tungsten carbide knives mounted on a Polycut-S motor-driven microtome (Reichert-Jung, Germany).2
Histological analyses showed for the first time that highly purified naturally-derived bovine osteogenic protein fractions set into motion the induction of cementogenesis with de novo induction of Sharpey's' fibers directly inserting into mineralized dentine (Figs. 7A, B, C) and within the newly forming cementoid matrix along the regenerated periodon-tal ligament system (Fig. 7D).2
Of interest, the insertion of Sharpey's fibers within the dentine and/or the cementoid matrix regulates the cellular trafficking close to the developing cementum whereby single fibers are directing staked progenitors towards the root planed surfaces for continuous cementoid induction (Figs. 7A, B, C, I magenta arrows).
The quality of the undecalcified sections allowed to identify histological features previously unreported such as the presence of condensations of cellular elements within the periodontal ligament space with condensed chromatin indicating de novo angiogenesis within the periodontal ligament space (Figs. 7B, C white arrows ).9,14,15
Capillaries were seen to be suspended by periodontal ligament fibers uniting the cementum to the basement membrane of the vessel (Fig. 7H). Fibers run from ce-mentum to the vessels as a conduit between the an-giogenic vascular and cementogenic microenvironments within the periodontal ligament system (Figs. 7H, I). "The role of the vessels in angiogenesis" is shown by an original Trueta' image34 (Fig. 7G). The electron microscopy image shows the exquisite relationships between osteoblasts (magenta arrow) and endothelial cells (light blue arrow). Cellular compartments are only separated by the ultrathin basement membrane with its extracellular matrix components including laminin and type IV collagen. 27,34,46,87
We also reported the presence of mesenchymal cells riding the fibers across the periodontal ligament space providing thus continuous progenitors cells to the forming cementum and cementoid matrices. Stacked cells are parked amongst the fibers against the root surface de novo initiating cementogenesis (Figs. 7H, I white arrows).1,9,17,18
The folding of the continuously erupting dental lamina of Carcharinus obscurus dusky shark shows pronounced immunolocalization of TGF-ß3 gene product (Figs. 6C. D).1 TGF-ß3 also immunolocalizes cellular elements below the folding of the dental lamina (Figs. 6C. D).1
This vast pleiotropic multi-functional poli-faceted biological activity of the TGF-ß3 gene and gene product across phylogenetically distant species and genera is additionally shown by the morphogenetic drive to induce substantial cementogenesis along planed root surfaces in Papio ursinus (Fig. 8). The molecular rational for the induction of cementogenesis is that research experiments have shown that TGF-ß signaling regulates cementum formation through Osterix expression.94 The experiments using conditional knockout mice for Osteocalcin and Tgfbr2 showed that TGF-ß signaling is one of the upstream regulators of Osterix in cementoblast differentiation and cementum formation.94
Osterix (Osx) is a key regulator of cellular cementum for-mation.94,95 Choi et al.94 demonstrated that a functional lack of Tgfbr2 determines a decrease in the expression of Osx thereby controlling the induction of cementogen-esis.94 TGF-ß signaling master minds cementoblasts differentiation by directly regulating Osx expression via a Smad-dependent pathway.94 The molecular complexities of the induction of cementogenesis are further highlighted by a study that shows that Osx-expressing cells express Wnt and that Wnts produced by Osx-activated cells regulate Wnts activated cementoblastic cells to proliferate and differentiate.96 Additional and novel molecular pathways have shown that Osx, an essential transcription factor for osteogenesis and cementogenesis, positively regulates DICKKORF-related protein (Dkk-1) to down-regulate the Wnt/ß-catenin pathway controlling osteoblasts and cementoblasts proliferation.97
The present experiments in P. ursinus' furcation defects super activated by 75 hTGF-ß3 in Matrigel®Matrix show up-regulation of Osteocalcin and Cementum protein-1 (Cemp-1). More importantly, and mechanistically determinant, the TGF-ß3 gene was up-regulated (Fig. 10).
Up-regulation of TGF-ß3 together with up-regulation of Osteocalcin and Cemp-1 resulted in the induction of substantial cementogenesis prominently forming along the planed and curetted root surfaces (Fig. 10).
The substantial induction of cementogenesis by the hTGF-ß3 isoform goes beyond the induction of cemento-genesis; 75 hTGF-ß3 in Matrigel®Matrix besides de novo inducing cementogenesis along the exposed root surfaces set the induction of collagenous conduits that molecularly and morphologically connect the sprouting capillaries to the cementoid microenvironment (Fig. 10).
Of interest, there was variable down-regulation of OP-1 and BMP-2 (Fig. 10). In context of periodontal tissue regeneration, the induction of alveolar bone and ceme-ntogenesis by the hTGF-ß3 isoform may not require up-regulation of selected BMPs genes for the induction of periodontal tissue regeneration, as in heterotopic induction of bone formation, constructing bone organoids as shown in Figure 2.11,58,61
In our studies in Class II furcation defects of the Chacma baboon Papio ursinus implanted with hTGF-ß3,19 there is induction of substantial cementogenesis along the root surfaces with deposition of cementoid matrix extending within the periodontal ligament space. It is noteworthy that newly formed cementum is penetrated by vascular canals, not only penetrating the as yet to be mineralized cementoid matrix but also in tight apposition to the newly forming cementoid matrix along the planed root surfaces (Fig. 8B). Implantation of hTGF-ß3 in Matrigel®Matrix show the substantial induction of both cementogene-sis and alveolar bone regeneration with the induction of periodontal ligament fibers within a highly vascularized periodontal ligament complex (Fig. 8).19
The induction of cementogenesis with capillary sprouting within the periodontal ligament space was also observed to a certain extent in control defects treated by extracellular matrix components in Matrigel®Matrix solo.19 Previous studies did show the morphogenetic inductive capacity of Matrigel Matrix™ solo in modulating the epithelial phenotype.87
Capillary sprouting was seen in close proximity of the newly formed cementum that however per se showed the presence of vascular canals within the newly forming cementoid matrix (Fig. 8). Identified vascular canals are seen within constructs attached to dentin matrices and penetrating the as yet to be mineralized cementoid (Fig. 8F magenta arrow). Capillaries penetrate the newly formed cementoid, surrounded by mineralized newly formed cementum (white arrows) (Fig. 8F).
High power morphological analyses show the previously unreported drive of vascular invasion, capillary sprouting and angiogenesis in constructing regenerating cemen-tum by doses of the recombinant hTGF-ß3 (Figs. 8,10). The unique and first morphological observations of vascular canals surrounded by cementoid matrix within mineralized cementum are shown in Figures 8 and 10. Close to the newly formed cementum in the periodontal ligament space there is a rich capillary sprouting and network almost attached to the cementoid matrix facing the periodontal ligament space (Figs. 8,10). This significant angiogenesis and capillary sprouting defines a novel term for the induction of cementogenesis, i.e. "ce-mentogenesis in angiogenesis" as we have previously defined "osteogenesis in angiogenesis".27,28,45
Of great interest, newly formed capillaries within the periodontal ligament (Fig. 8C,D,E) show a pattern of alignment and organization within the ligament space with fibers running from the vascular endothelial compartment to other capillaries or directly into the newly synthesized cementoid layer (Figs. 8D,E).
This morphological finding suggests that the newly formed capillaries within the periodontal ligament space provide a tri-dimensional construct geared to provide a biological structural passageway for both progenitors and soluble molecular signals from the vascular angiogenic compartment to the cementoid compartment along the root surface controlled by morphogenetic gradients across anatomical boundaries. Such boundaries include collagen fibres, endothelial basement membranes and the extra-cellular matrix with soluble morphogenetic signals in solution.
Nature thus constructs somehow simple passageways for movement across gradients of soluble morphogene-tic signals which may be both in solution or bound to extracellular matrix components including type IV collagen, entactin, nidogen and laminin, the whole ensemble constructing the induction of tissue morphogenesis.27,87
Finally, to further understand periodontal tissue induction and regeneration, we were able to implement one of the first in vivo studies correlating tissue induction and morphogenesis with molecular biology analyses of a set of morphogenetic genes controlling cementogenesis and the induction of alveolar bone in Class II furcation defects of Papio ursinus.19
Harvested tissues, i.e. cementum and alveolar bone on day 30 (whilst debriding the furcation defects created on day 0 of the experiments) and on day 90 (60 days after debridement and implantation of Matrigel®Matrix solo or 75pg of hTGF-ß3 in Matrigel®Matrix) were processed for quantitative reverse transcription polymerase chain reaction (qRT-PCR).19 Results showed induction of ce-mentogenesis by Matrigel®Matrix solo on days 60 after implantation of Matrigel®Matrix once again indicating the critical role of basement membranes component's in controlling tissue induction and morphogenesis.13,18,19,89 There was induction of angiogenesis (Figs. 9B,C,D) with up-regulation of BMP-2, TGF-ß3 and Osteocalcin particularly on day 90 with a two fold increase with respect to day 30 (Fig. 9A).
Control Matrigel®Matrix solo specimens showed down regulation of cementum protein-1 (Cempl) and up-regulation of TGF-ß3 on both periods (Fig. 9A).19 These results together with up-regulation of Osteocalcin also at both time periods may account for the cementogenic drive as shown morphologically by Matrigel®Matrix solo-treaded furcation defects (Figs. 9A,B).19 The data once again show the critical role of extracellular matrix components in the modulation of inductive phenomena as regulated by specific basement membrane components, particularly laminin and type IV.46,27,87
Implantation of Matrigel®Matrix reconstituted with 75 hTGF-ß3 showed downregulation of BMP-2 and OP-1 genes with up-regulation of TGF-ß3 and particularly of Osteocalcin both on days 30 and 9θ, with Cempl up-regulation between 30 and 90 days, the latter with a fourfold increase on day 90 as compared to day 30 (Fig. 10A). The reported gene expression profile (Fig. 10A)19 resulted in substantial tissue induction and morphogenesis as shown morphologically by the induction of cemento-genesis along the planed root surfaces 60 days after implantation of hTGF-ß3 (Figs. 8,10,C,D and inset A).
In ending, we would like to answer the question set at the very beginning of this communication: "Does the presence of multiple forms of BMPs have a therapeutic significance?" The selected second last iconographic plate of this review shows histological results outlining different morphological outcomes directly related to the specific amino acid sequence domain of each tested isoform singly or in binary application (Fig. 11 ). Molecularly, the presence of multiple forms of BMPs underlies the biological significance of apparent redundancy, and indicates multiple synergistic and synchronous interactions during tissue induction and morphogenesis beyond the realm of bone induction and regeneration.7,9,27
Implantation of highly purified osteogenic fractions after gel filtration chromatography onto tandem Sephacryl S-200 columns shows regeneration of alveolar bone (Fig. 11A), newly formed cementum with periodontal ligament fibres as collagenic bundles uniting the newly formed bone to the regenerated mineralized cementum with tightly inserted Sharpey's fibres (Fig. 11B).2
A carrier matrix of insoluble collagenous bone with highly purified osteogenic fractions purified greater than 60,000 fold with respect to the crude guanidinium extract2,27 has had a lesson to teach: optimal osteogenesis with the induction of cementogenesis with a functionally orientated periodontal ligament is the result of several genes and gene products expressed and secreted during regeneration that singly, synergistically and synchronously initiate, maintain and control tissue induction and morphogenesis.
We found that the original polyedric multifaceted mor-phogenetic drive of the intact bone matrix has all the required morphogens to set tissue induction and regeneration comparable with recombinant human proteins as shown by results comparing highly purified naturally derived osteogenic fractions to recombinant human os-teogenic proteins in non-human primate models.9,15,18
To potentiate the biological activity of hBMP-2 with limited cementogenic induction, a combination study with binary application of 100pg hOP-1 and 100pg hBMP-2 1:1 ratio by weight was implemented in Papio ursinus.8 The study showed that periodontal tissue induction and morphogenesis were qualitatively different when the morphogens were applied singly, with hOP-1 inducing substantial cementogenesis.
hBMP-2 treated defects showed, on the other hand, limited cementogenesis but a temporal enhancement of bone formation.8 Binary application showed cemento-genesis together with the induction of alveolar bone regeneration with marked osteoid synthesis (Fig. 11C).
In Class II furcation defects of Papio ursinus, binary application hOP-1/hTGF-ß3 20:1 ratio by weight showed substantial induction of periodontal tissue induction tempered however by the anatomy of the furcation model that could not hold the rapid induction of large ossicles within the treated furcation defects. Cementogenesis was substantial, and characterized by irregular thickened cemen-tum patches along the exposed and planed root surfaces (Fig. 11D).98
The biological acceptance of the inductive activity of a single recombinant human morphogen above the natural milieu and equilibrium of a pleiotropic bone matrix endowed with several naturally derived proteins clustered within the extra cellular matrix of bone has been the fundamental biological error of biotech companies developing recombinant hBMPs for translation in clinical contexts. Together with companies, far too eager clinician scientists embraced the powerful biological activity of a single recombinant human morphogen with no proper efficacy data beyond in vitro and in vivo rodent models' experimentation.27,28,61-63
Additional problems from the biotech industry were the firm decision to market single BMPs proteins as a recombinant human bone morphogenetic protein without consideration whatever to the structure/activity profile, possibly because of the impending FDA regulations on approving or not approving osteogenic proteins and certainly a single recombinant human protein rather than a combination thereof, mimicking the multiple morpho-genetic capacity of purified chaotropically extracted bone matrices.
A final, and as it turned out, lethal error of biotech companies, was to seek approval for much higher doses than the doses used and filed in pre-clinical animal studies. Aside the vague rational that higher doses were needed in clinical contexts, FDA approval was requested for much higher doses of recombinant proteins than the doses used in pre-clinical studies including non-human primates to initiate human osteoinduction. It turned out that even massive dose of several milligram proteins per gram of carrier were needed to yield insufficient regeneration often inferior to autogenous bone grafts.28,45,91
The induction of substantial cementogenesis in angio-genesis as observed in hTGF-ß3-treated specimens but particularly the induction of large trabeculated constructs along the planed root surface indicates that the TGF-ß 3gene and gene product are recapitulating in extant Papio ursinus the induction of trabeculated cementum with vascular canals as observed in extinct Mosasaurs, 168-165 Ma (Fig. 12).1,99 It is likely that the TGF-ß3 gene and gene product have been deployed and re-deployed during phylogenetically ancient evolutionary pathways controlling the induction of mineralized matrices including cementum across distant phyla (Fig. 12).
Acknowledgments
The University of the Witwatersrand, Johannesburg and the South African National Research Foundation have supported our multiple studies on the recombinant hTGF-ß3. I thank Novartis AG for supplying the recombinant mor-phogen. A special thanks to the molecular biology team headed by Raquel Duarte in the School of Clinical Medicine - Internal Medicine for together cracking the molecular insights of the induction of bone formation by the recombinant hTGF-ß3 in Papio ursinus. This work could not have been possible without the unique expertise of Barbara van den Heever in cutting undecalcified blocks of dentine, cementum and alveolar bone providing the first morphological insights into the induction of cementogenesis in Papio ursinus. A special thanks to Ruqayya Parak for having added to the Bone Research Unit the new dimension of her unique expertise in cutting undecalcified sections on the Exakt diamond saw and grinding equipment, donated to the Unit by a Wellcome Trust grant in 1998.
References
1. Ripamonti U. Developmental pathways of periodontal tissue regeneration. Developmental diversities of tooth morphogenesis do also map capacity for periodontal tissue regeneration? J Periodont Res. 2019; 54(1), 10-26. https://doi:10.1111/jre.12596. Epub 2018 Sep 12. [ Links ]
2. Ripamonti U, Heliotis M, van den Heeer B,Reddi AH. Bone morphognetic proteins induce periodontal regeneration in the baboon (Papio ursinus). J Periodont Res, 1994; 29(6), 439-45. https://doi:10.1111/j.1600-0765.1994.tb01246.x. [ Links ]
3. Ripamonti U, Reddi AH. Periodontal regeneration: potential role of bone morphogenetic proteins. J Periodont Res, 29(4), 225-35. https://doi:10.1111/j.1600-0765.1994.tb01216.x. [ Links ]
4. Ripamonti U. Induction of cementogenesis and periodontal ligament regeneration by bone morphogenetic proteins. In Bone Morphogenetic Proteins: Biological Characteristics and Reconstructive Repair. TS Lindholm, (Ed.) RG Landes Company and Academic Press, Inc., 1996; (17). 189-98. [ Links ]
5. Ripamonti U, Heliotis M, Sampath TK, Rueger D. Induction of cementogenesis by recombinant human osteogenic pro-tein-1 (hOP-1/BMP-7) in the baboon (Papio ursinus). Arch Oral Biol, 1996; 41(1), 121-6. https:/doi:10.1016/0003-9969(95)00110-7. [ Links ]
6. Ripamonti U, Reddi AH. Tissue engineering, morphogenesis and regeneration of periodontal tissues by bone morphoge-netic proteins. Crit Rev Oral Biol Med, 1997; 8(2), 154-63. https://doi:10.1177/10454411970080020401. [ Links ]
7. Thomadakis G, Ramoshebi, LN, Crooks J, Rueger DC, Ripa-monti U. Immunolocalization of bone morphogenetic protein -2 and -3 and osteogenic protein-1 during murine tooth root morphogenesis and in other craniofacial structures. European. Journal of Oral Sciences, 1999; 107(5), 368-77. https://doi:10.1046/j.09098836.1999.eos107508.x. [ Links ]
8. Ripamonti U, Crooks J, Petit J-C, Rueger DC. Periodontal tissue regeneration by combined applications of recombinant human osteogenic protein-1 and bone morphogenetic pro-tein-2. A pilot study in Chacma baboons (Papio ursinus). European. Journal of Oral Sciences, 2001; 109(4), 241-8. https://doi:10.1034/j.16000722.2001.00041.x. [ Links ]
9. Ripamonti U. Recapitulating development: a template for peri-odontal tissue engineering. Tissue Engineering. 2007; 13(1), 51-71. https://doi:10.1089/ten.2006.0167. [ Links ]
10. Teare JA, Ramoshebi LN, Ripamonti U. Periodontal tissue regeneration by recombinant human transforming growth factor -ß3in Papio ursinus. Journal of Periodontal Research, 2008; 43(1), 1-8. https://doi:10.1111/j.1600-0765.2007.00987.x. [ Links ]
11. Ripamonti U, Ramoshebi LN, Teare J, Renton L, Ferretti C. The induction of endochondral bone formation by transforming growth factor-ß3: Experimental studies in the non-human primate Papio ursinus. J Cell Mol Med. 2008; 12(3), 1029-48. https://doi:10.1111/j.1582-4934.2008.00126.x. [ Links ]
12. Ripamonti U, Teare J, Petit J-C. Induction of cementogenesis and periodontal ligament by bone morphogenetic proteins. In S Vukicevic (Ed.), Bone Morphogenetic Proteins: From Local to Systemic Therapeutics. Birkauser Verlag AG, Basel/ Switzerland. 2008; 233-56. [ Links ]
13. Ripamonti U, Parak R, Petit J-C. Induction of cementogene-sis and periodontal ligament regeneration by recombinant human transforming growth factor-ß3 in Matrigel with rectus abdominis responding cells. J Periodont Res. 2009; 44(1), 141-52. https://doi:10.1111/j.1600-0765.2008.01086.x. [ Links ]
14. Ripamonti U, Petit J-C, Teare J. Cementogenesis and the induction of periodontal tissue regeneration by the osteo-genic proteins of the transforming growth factor-ß superfamily. J Periodont Res. 2009; 44(2): 81-7. https://doi.org/10.1111/j.1600-0765.2008.01158.x. [ Links ]
15. Ripamonti U, Petit J-C. Bone morphogenetic proteins, ce-mentogenesis, myoblastic stem cells and the induction of periodontal tissue regeneration. Cyt Growth Factor Rev. 2009; 20 (5-6), 489-99. https://doi.org/10.1016/jcytogfr.2009.10.016 [ Links ]
16. Ripamonti U, Klar M, Renton LF, Ferretti C. Synergistic induction of bone formation by hOP-1, hTGF-ß3 and inhibition by zoledronate in macroporous coral derived hydroxyapatites. Biomaterials, 2010; 31(25): 6400-10. https://doi:10.1016/j.biomaterials.2010.04.037. Epub 2010 May 21. [ Links ]
17. Ripamonti U, Petit J-C, Teare J. Tissue Engineering of the Periodontal Tissues In: Regenerative Dentistry, MK Marei and KA Athanasiou (Eds) Morgan & Claypool Publishers. 2010; (3): 83-109. [ Links ]
18. Ripamonti U. Redefining the induction of periodontal tissue regeneration in primates by the osteogenic proteins of the transforming growth factor-ß supergene family. J Periodont Res, 2016; 51(6): 699-715. https://doi:10.1111/jre.12356.Epub Feb 2. [ Links ]
19. Ripamonti U, Parak R, Klar RM, Dickens C, Dix-Peek T, Duarte R. Cementogenesis and osteogenesis in periodontal tissue regeneration by recombinant human transforming growth factor-ß3: a pilot study in Papio ursinus. J Clin Periodontol, 2017; 44(1), 83-95. https://doi:10.1111/jcpe.12642. Epub 2016 Dec 1. [ Links ]
20. Levander G. Tissue induction. Nature. 1945; 155: 148-9. https://doi.org/10.1038/155148a0. [ Links ]
21. Urist MR. Bone: formation by autoinduction. Science. 1965; 150(3698): 893,899,220,680-6. http://doi:10.1126/science.150.3698.893. [ Links ]
22. Sampath TK, Reddi AH. Dissociative extraction and reconstitution of extracllular matrix components involved in local bone differentiation. Proc Natl Acad Sci USA, 1981; 78(12): 7599-603. https://doi.1073/pnas.78.12.7599. [ Links ]
23. Sampath TK, Reddi AH. Homology of bone inductive proteins from human, monkey, bovine, and rat extracellular matrix. Proc Natl Acad Sci USA, 1983; 80(21): 6591-95. https://doi:10.1073/pnas.80.21.6591 [ Links ]
24. Reddi AH. Bone morphogenesis and modeling: soluble siganls sculpt osteosomes in the solid state. 1997; Cell, 89(2): 159-61. https://doi:10.1016/s0092-8674(00)80193-2. [ Links ]
25. Urist MR, Silverman BF, Büring K, Dubuc FL, Rosenberg JM. The bone induction principle. Clin Orthop Rel Res, 1967; 53: 243-83. [ Links ]
26. Ripamonti U. Osteogenic proteins of the transforming growth factor-ß superfamily. In: HL Henry and AW Norman (Eds.), Encyclopedia of Hormones. Academic Press, 2003; 80-6. [ Links ]
27. Ripamonti U. Soluble osteogenic molecular signals and the induction of bone formation. Biomaterials, 2006; 27(6): 807-22. https://doi:10.1016/j.biomaterials.2005.09.021. Epub 2005 Oct5. [ Links ]
28. Ripamonti U, Ferrett, C, Heliotis M. Soluble and insoluble signals and the induction of bone formation: Molecular therapeutics recapitulating development. J Anat, 2006; 209(4): 447-68. https://doi:10.1111/j.1469-7580.2006.00635.x. [ Links ]
29. Kilkenny C, Browne WJ,Cuthill IC, Emerson M, Altman DG. Improving bioscience research reporting the ARRIVE guidelines for reporting animal research. PLoS Biol. 2010; 8, c100 0412. [ Links ]
30. Turing AM. The chemical basis of morphogenesis. Phil Transact Royal Soc London. Series B, Biol Sci. 1952; B237(641): 37-72. [ Links ]
31. Ripamonti U. Functionalized surface geometries induce "Bone: Formation by autoinduction". Front Physiol. 2018; 8: 1084 https://doi:10.3389/fphys.2017.01084. [ Links ]
32. Sacerdotti C, Frattin G. Sulla produzione eteroplastica dell' osso. Rivista Accademica Medica, Torino. 1901; 27: 825-36. [ Links ]
33. Huggins CB. The formation of bone under the influence of epithelium of the urinary tract. Arch Surg. 1931; 22: 377-408. [ Links ]
34. Trueta J. The role of vessels in osteogenesis. J Bone Joint Surg. 1963; 45B: 402-18. [ Links ]
35. von Haller A. Experimentorium de ossium formatione, In Opera minora, Vol. 2, Grasset F, Ed. Lausanne. 1973; 400. [ Links ]
36. Lanza D, Vegetti M. Opere biologiche di Aristotele. A cura di Diego Lanza e Mario Vegetti, UTET, Torino. 1971. [ Links ]
37. Crivellato E, Nico B, Ribatti D. Contribution of endothelial cells to organognesis: a modern reappraisal of an old Aristotelian concept. J Anatomy. 2007; 211 (4): 415-27. https://doi:10.1111/j.1469-7580.2007.00790.x. Epub 2007 Aug 7. [ Links ]
38. Gomez-Salinero JM, Rafii S. Endothelial cell adaptation in regeneration. Science. 2018; 362(419): 1116-11. https://doi:10.1126/science.aar4800. [ Links ]
39. Senn N. On the healing of aseptic bone cavities by implantation of antiseptic decalcified bone. Am J Med Sci, 1889; 98: 219-43. https://doi:10.1097/00000658-188907000-00043. [ Links ]
40. Reddi AH. Morphogenesis and tissue engineering of bone and cartilage: inductive signals, stem cells, and biomimetic biomaterials. Tissue Eng. 2000; 6(4): 351-59. https://doi:10.1089/107632700418074. [ Links ]
41. Levander G. A study of bone regeneration. Surg Gynec Obst, 1938; 67(6): 705-14. [ Links ]
42. Levander G, Willestaedt H. Alcohol-soluble osteogenetic substance from bone marrow. Nature, 1946; 4,157: 87. https://doi:10.1038/157587b0. [ Links ]
43. Moss ML. Extraction of an osteogenic inductor factor from bone. Science. 1958; 127(3301): 755-6. https://doi:10.1126/ science.127.3301.755. [ Links ]
44. Reddi AH, Huggins CB. Biochemical sequences in the transformation of normal fibroblast in adolescent rats. Proc Natl Acad Sci USA, 1972; 69(6): 1601-5. https://doi:10.1073/pnas.69.6.1601. [ Links ]
45. Ripamonti U, Heliotis M, Ferretti C. Bone morphogenetic proteins and the induction of bone formation: From laboratory to patients. Oral Maxfac Surg.Clin of North Am, 2007; 19(4): 575-89. https://doi:10.1016/j.coms.2007.07.006. [ Links ]
46. Vukicevic S, Luyten FP, Kleinman HK, Reddi AH. Differentiation of canalicular cell processes in bone cells by basement membrane matrix components: regulation by discrete domains of laminin. Cell. 1990; 63(2) 437-45. https://doi:10.1016/0092-8674(90)90176-f. [ Links ]
47. Kusumbe AP, Ramasamy SK, Adams RH. Coupling of angio-genesis and osteogenesis by a specific vessel subtype in bone. Nature. 2014; 507: 323-8. https://doi:10.1038/nature 13145. Epub 2014 Mar 12. [ Links ]
48. Ramasamy SK, Kusumbe AP, Adams RH. Regulation of tissue morphogenesis by endothelial cell-derived signals. Trends in Cell Biol. 2015; 25(3): 148-57. https://doi:10.1016/j.tcb. 2014.11.007. Epub 2014 Dec 17. [ Links ]
49. Medici D, Shore EM, Lounev VY, Kaplan FS, Kalluri R. Conversion of vascular endothelail cells into multipotent stem-like cells. Nature Med. 2010; 16(12): 1400-06. https://doi:10.1038/nm.2252. Epub 2010 Nov 21. [ Links ]
50. Ramasamy SK, Kusumbe, AP, Wang L, Adams RH. Endothe-lial Notch activity promotes angiogenesis and osteogenesis in bone. Nature. 2014; 507(7492): 376-80. https://doi:10.1038/nature13146. Epub 2014 Mar 12. [ Links ]
51. Heliotis M, Ripamonti U. Phenotypic modulation of endothelial cells by bone morphogenetic protein fractions in vitro. In Vitro Cell Develop Biol, 1994; 30A(6): 353-5. https://doi:10.1007/BF02634354. [ Links ]
52. Leversha M, Brennan C,Tabar V. Glioblastoma stem-like cells give rise to tumour endothelium. Nature. 2010; 468(7325), 829-33. https://doi:10.1038/nature09624. Epub 2010 Nov 21. [ Links ]
53. Ripamonti U, Parak R, Klar MR, Dickens C, Dix-Peek T, Duarte R. The synergistic induction of bone formation by the osteogenic proteins of the TGF-ß supergene family. Biomaterials. 2016; 104: 279-96. https://doi:10.1016/j.biomaterials.2016.07.018. Epub 2016 Jul 20. [ Links ]
54. Ripamonti U, Duneas N, van den Heever B, Bosch C, Crooks J. Recombinant transforming growth factor-ß1 induces endo-chondral bone in the baboon and synergizes with recombinant osteogenic protein-1 (bone morphogenetic protein-7) to initiate rapid bone formation. J Bone Miner Res, 1997; 12(10): 1584-95. https://doi:10.1359/jbmr.1997.12.10.1584. [ Links ]
55. Klar RM, Duarte R, Dix-Peek T, Ripamonti U. The induction of bone formation by the recombinant human transforming growth factor-ß3. Biomaterials. 2014; 35(9), 2773-88. https://doi:10.1016/j.biomaterials.2013.12.062. Epub 2014 Jan 15. [ Links ]
56. Ripamonti U. Osteogenic device for inducing bone formation in clinical contexts. US 2012/0277879 A1. 2012; Publication date 1 November 2012. [ Links ]
57. Ripamonti, U. Osteogenic device for inducing bone formation in clinical contexts. US 9,084,757 B2. 2015; Publication date 21 July 2015. [ Links ]
58. Ripamonti, U. Induction of Bone Formation in Primates. The Transforming Growth Factor beta 3, CRC Press, Taylor & Francis Group, Boca Raton, USA, 2016. [ Links ]
59. Massagué J, Blain W, Lo RS. TGFbeta signaling in growth control, cancer, and heritable disorders. Cell. 2000; 103(2): 295-309. https://doi:10.1016/s0092-8674(00)00121-5. [ Links ]
60. Kicheva A, González-Gaitán M. The Decapentaplegic mor-phogen gradient: a precise definition. Curr Opin Cell Biol, 2008; 20(2), 137-43. https://doi:10.1016/j.ceb.2008.01.008. [ Links ]
61. Ripamonti U, Dix-Peek T, Parak R, Milner B, Duarte R. Profiling bone morphogenetic proteins and transforming growth factor-ßs by hTGF-ß3 pre-treated coral derived macroporous constructs: The power of one. Biomaterials, 2015; 49: 90-102. https://doi:10.1016/j.biomaterials.2015.01.058. Epub 2015 Feb 14. [ Links ]
62. Ripamonti U, Duarte R, Ferretti C. Re-evaluating the induction of bone formation in primates. Biomaterials, 2014; 35: 9407-22. https://doi:10.1016/j.biomaterials.2014.07.053. Epub 2014 Aug 23. [ Links ]
63. Ripamonti U, Duarte R, Parak R, Dickens C, Dix-Peek T, Klar RM. Redundancy and molecular evolution: The rapid induction of bone formation by the mammalian transforming growth factor-ß3 isoform. Front Physiol, 2016; 7, 396. https://doi:10.3389/fphys.2016.00396. eCollection 2016. [ Links ]
64. Shi Y, Massagué J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell. 113(6): 685-700. https://doi:10.1016/s0092-8674(03)00432-x. [ Links ]
65. Feng XH, Derynck R. (2005). Specificity and versatility in tgf-beta signaling through. Smads. Annual Rev Cell Dev Biol, 2005; 21: 659-93. https://doi:10.1146/annurev.cellbio.21.022 404.142018. [ Links ]
66. Massagué J, Seoane J, Wotton D. Smad transcription factors. Genes Develop, 2005; 19(23): 2783-810. https://doi:10.1101/gad.1350705. [ Links ]
67. Ross S, Hill CS. How the Smads regulate transcription. Int J Biochem Cell Biol. 2008; 40(3): 383-408. https://doi:10.1016/j.biocel.2007.09.006. Epub 2007 Oct 7. [ Links ]
68. Mullen AC, Orlando A, Newman JJ, Lovén J, Kumar RM, Bilodeau S, Reddy J, Guenther MG, DeKoter RP, Young RA. Master transcription factors determine cell-type-specific responses to TGF-ß signaling. Cell. 2001; 147(3): 565-76. https://doi:10.1016/j.cell.2011.08.050. [ Links ]
69. Massagué J, Wotton D. Transcriptional control by the TGF-beta/Smad signaling system. EMBO J. 2000; 19(8): 1745-54. https://doi:10.1093/emboj/19.8.1745. [ Links ]
70. Lee KS, Hong SH, Bae S.C. Both the Smad and p38 MAPK pathways play a crucial role in Runx2 expression following induction by transforming growth factor-beta and bone mor-phogenetic protein. Oncogene, 2002; 21(47): 7156-63. https://doi:10.1038/sj.onc.1205937. [ Links ]
71. Derynck R, Zhang YE. Smad-dependent and mad-independent pathways in TGF-beta Family signaling. Nature, 2003; 425 (6958), 577-84. https://doi:10.1038/nature02006. [ Links ]
72. Phinney DG, Prockop DJ. Concise review: mesenchymal stem/multipotent stromal cells: the state of transdifferentiation and modes of tissue repair - current views. Stem Cells. 2007; 25(11): 2896-902. https://doi:10.1634/stemcells.2007-0637. Epub 2007 Sep 27 [ Links ]
73. Seo B-M, Miura M, Gronthos S, et al. Investigation of multipotent postnatal stem cells. From human periodontal ligament. Lancet, 2004; 364(9429): 149-55. https://doi:10.1016/S0140-6736(04)16627-0. [ Links ]
74. Lin NH, Menicanin D, Mrozik K, Gronthos S, Bartold PM. Putative stem cells in regenerating human periodontium. J Periodont Res, 2008; 43(5), 514-23. https://doi:10.1111/j.1600-0765.2007.01061.x. [ Links ]
75. Jian H, Shen X, Liu I, Semenov M, He X, Wang X-F. Smad 3-dependent nuclear translocation of ß-catenin is required for TGF-ß1-induced proliferation of bone marrow derived adult human mesenchymal stem cells. Genes & Dev. 2006; 20: 666-74. [ Links ]
76. Crisan M, Yap S, Casteilla L, et al. A perivascular origin for mesenchymal stem cells in multiple human organs. Cell Stem Cell, 2008; 3(3), 301-13. https://doi:10.1016/j.stem.2008.07.003. [ Links ]
77. Wang X, Harris RE, Bayston LJ, Ashe HL. Type IV collagens regulate BMP signalling in Drosophila. Nature, 2008; 455 (7209): 72-77. https://doi:10.1038/nature07214. [ Links ]
78. Sampath TK, Raska KE, Doctor JS, Tucker RF, Hoffmann FM. Drosophila transforming growth factor beta superfamily proteins induce endochondral bone formation in mammals. Proc Natl Acad Sci USA, 1993; 90(13): 6004-08. https://doi:10.1073/pnas.90.13.6004. [ Links ]
79. Roberts AB, Sporn MB, Assoian RK, et al. Transforming growth factor type beta: Rapid induction of fibrosis and an-giogenesis in vivo and stimulation of collagen formation in vitro. Proc Natl Acad Sci USA, 1986; 83(12): 4167-71. https://doi:10.1073/pnas.83.12.4167. [ Links ]
80. Assoian RK, Komoriva A, Meyers CA, Miller DN, Sporn MB. Transforming growth factor-beta in human platelets. Identification of a major storage site, purificaton, and characterization. J Biol Chem. 1983; 258(11): 7155-60. [ Links ]
81. Robey, PG, Young MF, Flanders KC, Roche NS, Kondaiah P, Reddi AH, Termine JD, Sporn MB, Roberts AB. Osteoblasts synthetize and respond totransforming growth factor-type beta (TGF-beta) in vitro. J Cell Biol. 1987; 105(1): 457-63. https://doi:10.1083/jcb.105.1.457. [ Links ]
82. Centrella M, Massagué J, Canalis E. Human platelet-derived transforming growth factor-ß stimulates parameters of bone growth in fetal rat calvaria. Endocrinology, 1986; 119(5), 2306 -12. https://doi:10.1210/endo-119-5-2306. [ Links ]
83. Noda J, Camilliere JJ. In vivo stimulation of bone formation by transforming growth factor-ß. Endocrinology, 1989; 124(6), 2991-4. https://doi:10.1210/endo-124-6-2991. [ Links ]
84. Joyce ME, Roberts AB, Sporn MB, Bolander ME. Transforming growth factor-ß and the initiation of chondrogenesis and osteogenesis in the rat femur. J Cell Biol, 1990; 110(6): 2195-2207. https://doi:10.1083/jcb.110.6.2195. [ Links ]
85. Seeherman HJ, Berasi SP, Brown CT, Martinez RX, Juo ZS, Jelinsky S, Cain MJ, Grode J, Tumelty KE, Bohner M, et al. A BMP/activin A chimera is superior to native BMPs and induces bone in nonhuman primates when delivered in a composite matrix. Science Translational Medicine. 2019; 11(489): eaar4953. https://doi:10.1126/scitranslmed.aar4953. [ Links ]
86. Ripamonti U, Duarte R. Tissue Transfiguration in vivo. South African Provisional Patent Spec. 2019/08010, Dec. 3, 2019. [ Links ]
87. Ripamonti U, van den Heever B, Heliotis M, Dal Mas I, Hahn-le U, Biscardi A. Local delivery of bone morphogenetic proteins using a reconstituted basement membrane gel: Tissue engineering with Matrigel. 2002; South Afr J Sci, 2002; 98 (9-10), 429-33. [ Links ]
88. Fatehullah A, Tan SH, Barker N. Organoids as an in vitro model of development and disease. Nature Cell Biol, 2016; 18(3): 246-54. https://doi:10.1038/ncb3312. [ Links ]
89. Balooch G, Balooch M, Nalla RK, Schilling S, Filvaroff EH, Marshall G., Marshall SJ, Ritchie RO, Derynk, R, Alliston T. TGF-ß regulates the mechanical properties and composition of the bone matrix. Proc Natl Acad Sci USA. 2005; 102(52): 18813-18. https://doi:10.1073/pnas.0507417102. Epub 2005 Dec 14. [ Links ]
90. Ripamonti U, Crooks J, Matsaba T, Tasker T. Induction of endochondral bone formation by recombinant human transforming growth factor-ß2 in the baboon (Papio ursinus). Growth Factors. 2000; 17(4): 269-85. https://doi:10.3109/08977190009028971. [ Links ]
91. Ferretti C, Ripamonti U. Long-term follow-up of pediatric mandibular reconstruction with human transforming growth factor-ß3. J Craniofac Surg. 2020; 31(5), 1424-29. https://doi:10.1097/SCS.0000000000006568. [ Links ]
92. Helder MN, Kar H, Bervoets TJM, Vukicevic S, Burger EH, D'Souza RN, Wöltgens JHM, Karsenty G, Bronkers ALJJ. Bone morphogenetic proteion-7 (Osteogenic Protein-1, OP-1) and tooth developent. J Dent Res, 1998; 77(4), 545-54. https://doi:10.1177/00220345980770040701. [ Links ]
93. Ripamonti U, Vukicevic S. Bone morphogenetic proteins: from developemntal biology to molecular therapeutics. South Afr J Sci. 1995; 91(6), 277-80. [ Links ]
94. Choi H, Ahn Y-H, Kim T-H, Bae C-H, Lee J-C, You H-K, Cho E-S (2016). TGF-β signaling regulates cementum formation through Osterix expression. Sci Rep. 2016; 6, 26046; https://doi:10.1038/srep26046(2016). [ Links ]
95. Cao Z, Liu R, Zhang H, Liao H, Zhang Y, Hinton RJ, Feng JQ. Osterix controls cementoblasts differentiation through down-regulation of Wnt-signaling via enhancing DKK1 expression. Int J Biol Sci. 2015; 11(3): 335-44. Doi:10.7150/ijbs.10874. [ Links ]
96. Tan SH, Senarath-Yapa K, Chung MT, Longaker MT, Wu JY, Nusee R. Wnts produced by osterix-expressing osteolineage cells regulate their proliferation and differentiation. Proc Natl Acad Sci USA. 2014; E5262-E5271. [ Links ]
97. Wang C, Liao H, Sun H, Zhang Y, Cao Z. MicroRNA-3064-3p regulates the differentiation of cementobslasts through target ing DKK1. J Periodont Res. 2018; 53: 705-13. [ Links ]
98. Teare JA, Petit J-C, Ripamonti U. Synergistic induction of peri-odontal tissue regeneration by binary application of human osteogenic protein-1 and human transforming growth factor-ß3 in Class II furcation defects of Papio ursinus. J Periodont Res, 2012; 47(3): 336-44. https://doi:10.1111/J.1600-0765.2011.01438.x. [ Links ]
99. Luan X, Walker C, Dangaria S, et al. The mosasaur tooth attachment apparatus as paradigm for the evolution of the gnathostome periodontium. Evol Develop. 2009; 11(3): 247-59 .https://doi10.1111/j.1525-142X.2009.00327.x. [ Links ]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
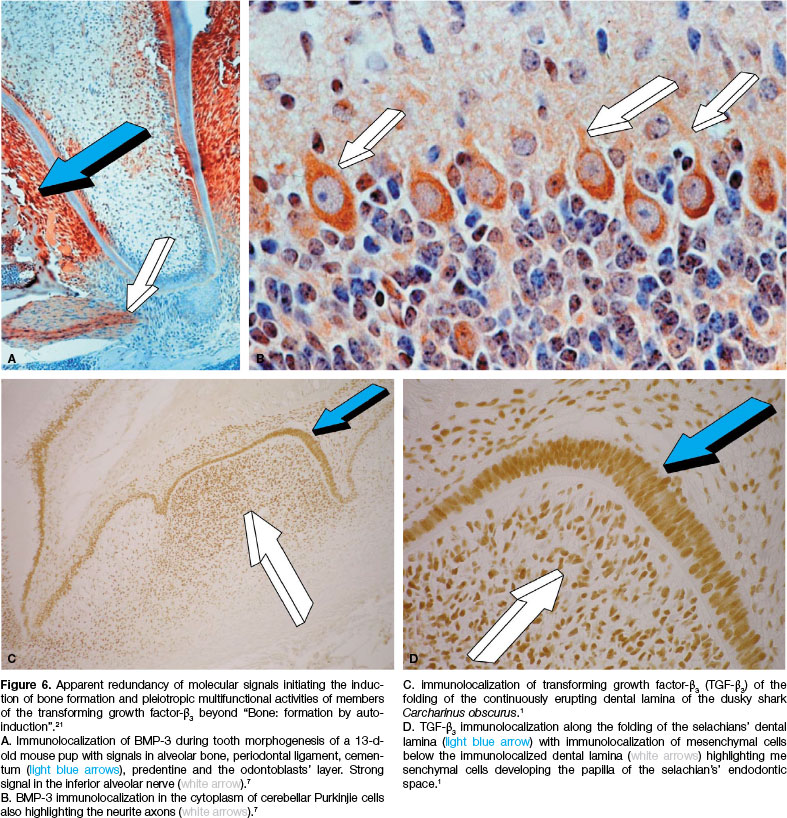{kind=link}
{kind=link}
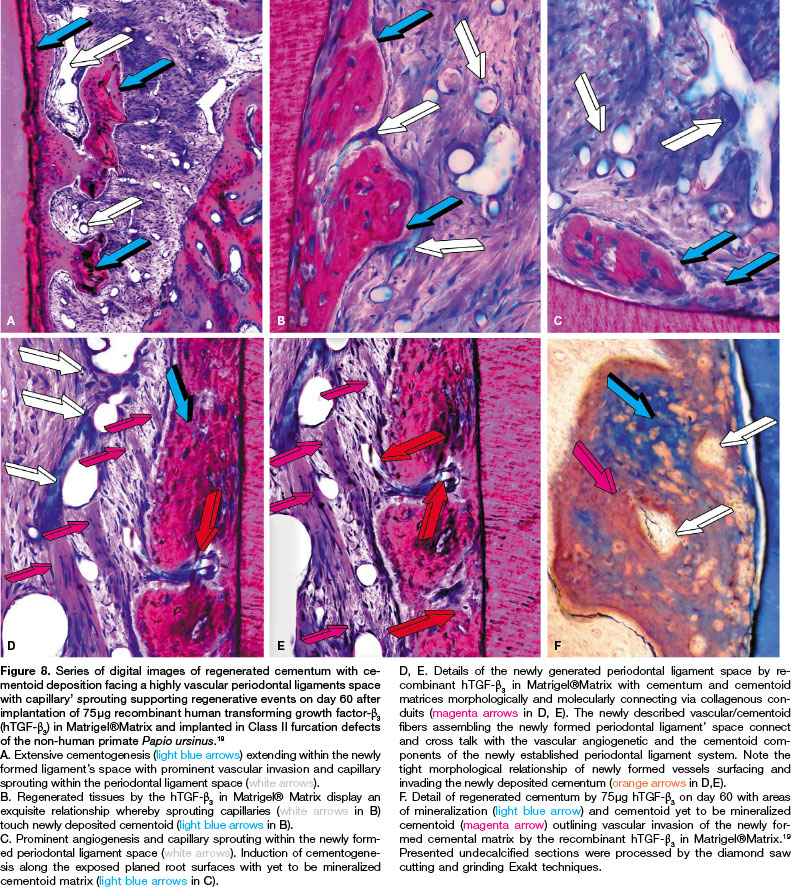{kind=link}
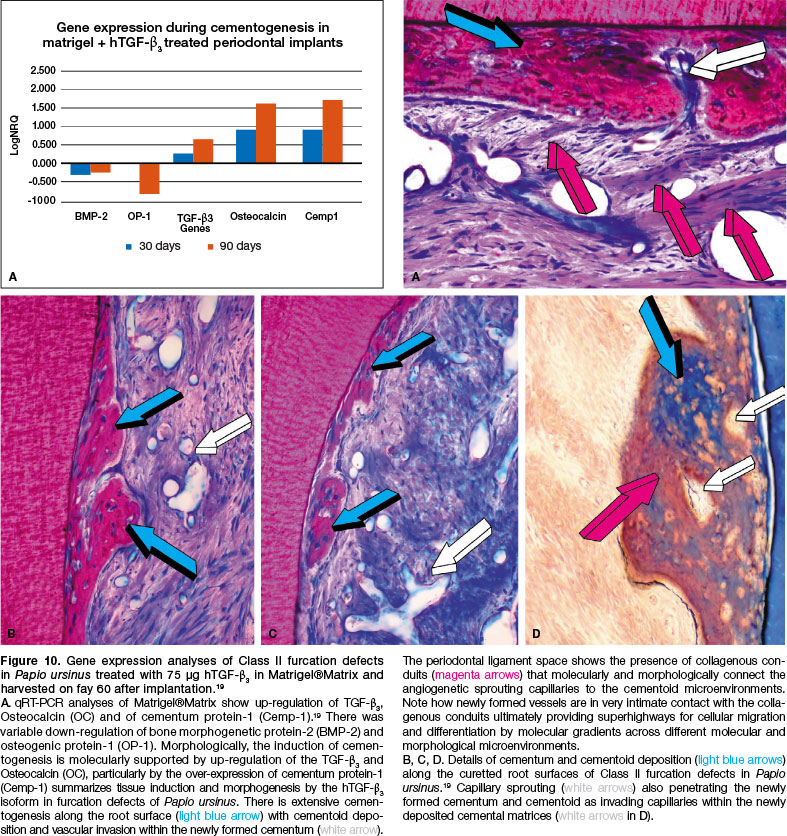{kind=link}
{kind=link}
{kind=link}
{kind=link}