










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.51 n.2 Pretoria 2021
http://dx.doi.org/10.38201/btha.abc.v51.i2.2
ORIGINAL RESEARCH
Habitat description of the rare orchid Didymoplexis verrucosa for more effective conservation
T.H.C. MostertI; R.E. MostertII
IBotany Dept., University of Zululand, KwaDlangezwa Campus, South Africa / Private Bag X1001, KwaDlangezwa, 3886
IILife Science, Felixton College, Felixton, South Africa / P.O. Box 252, Felixton, KwaZulu-Natal, 3875
ABSTRACT
BACKGROUND: Didymoplexis verrucosa is a cryptic, leafless, saprophytic ground orchid (~70 mm tall) growing on the coastal forest floors of southern Zululand and classified as Vulnerable (D2). As part of a population monitoring programme, 960 man-hours of species-specific searching over five consecutive flowering seasons were conducted, yielding only one individual plant
OBJECTIVE: The aim of this study was to increase detection rate by developing a profile of environmental indicators for the accurate identification of suitable habitat
METHODS: A detailed description of suitable habitat was compiled based on the Braun-Blanquet approach
RESULTS: The results showed that key attributes shared by localities include similar topographic position in the landscape, hydrology, soils, vegetation composition and structure, forest age, leaf-litter composition of the forest floor, the co-occurrence of Isoglossa woodii, and a similar degree of protection from sunlight, wind and desiccation
CONCLUSION: This profile of essential habitat characteristics can be used as a surrogate in the absence of actual locality data when identifying target conservation areas and compiling management strategies for this very cryptic species. A by-product of this habitat analysis was the discovery of a long list of impacts on the long term survival of D. verrucosa. The combination of these stochastic and deterministic events will drive habitat change at rates beyond the species' ability to adapt. Managing these variables forms the crux of its successful conservation. A revision of the conservation status, based on the formal IUCN criteria, indicates that D. verrucosa should be reclassified as Critically Endangered Category B2a and D
Keywords: habitat description, habitat fragmentation and degradation, Red Data List, saprophyte.
Introduction
Didymoplexis verrucosa J.Stewart & Hennessy (Orchidaceae: Epidendroideae) is a small and cryptic, leafless, saprophytic ground orchid discovery by the Dutch botanist C.G.G.J. Van Steenis in 1975 (Stewart & Hennessey 1980). It grows along the forest floors of the coastal dune cordon of southern Zululand (northern KwaZulu-Natal, South Africa). The known distribution range is restricted to the forests south and east of the town Mtunzini. These saprophytes obtain all the energy they need from parasitising the network of fungal hyphae within the leaf-litter carpet of the deeply shaded forest floors. They are therefore classified as mycoheterotrophs (Miura et al. 2018) and do not photosynthesise their own carbohydrates like most plants. This parasitic dependency on fungi evolved from the normally mutualistic relationship between plants and mycorrhiza fungi, which may have evolved many times over, independently in many different plant families (Miura et al. 2018).
Didymoplexus verrucosa forms small tubers (up to 90 x 20 mm) within the decomposed organic matter of the forest floor, accumulating resources from fungal hyphae around their root system (Stewart & Hennessey 1980). With no need to synthesise their own food, they do not invest in leaves and chlorophyll for photosynthesis. The accumulated resources are used in early spring to produce a single erect, very delicate, leafless, cream-coloured flowering stem (± 70 mm tall) bearing eight to twelve very small (10-13 mm in diameter) white flowers tinged pinkish brown (Stewart & Hennessey 1980). The flowers open sequentially, but each flower remains open for only a few hours for pollination before it closes again, and no floral odour has been detected (Stewart & Hennessey 1980). Cross pollination has not yet been recorded and the specific pollinating vector is still unknown. In the absence of insect pollination, Stewart and Hennessey (1980) observed potential cleistoga-mous self-pollination behaviour of the flowers.
Historical perspective of discovery and rarity
The search for D. verrucosa began long before its discovery. During the 1920s and 1930s Van Steenis compiled keys to the many saprophytic species of a wide range of flowering plants occurring in the tropical forests of the Malesia Floristic Region (Kalkman 1990). Based on the knowledge that he gained regarding suitable habitat for such species, he predicted that the coastal dune forests of Zululand would contain saprophytic orchid species (Stewart & Hennessey 1980). After numerous botanical expeditions to these Zululand forests, his wife found such a species, which he concluded belonged to the genus Didymoplexis. Based on the warty protrusions on the fruit capsule, it was named D. verrucosa (Stewart & Hennessey 1980). Initially, only a single plant was found on the Farm Twinstreams, then owned by conservationist Ian Garland. Subsequent searches in the following years revealed a small population numbering approximately 15 plants. Local ecologist C.J. Ward photographed the flowers for the first time in August 1978 and Garland continued to monitor the population for a few years. Subsequent recordings of these very cryptic plants became more sporadic, often with decades passing between sightings (Victor et al. 2005).
In the early 2000s, orchid enthusiast Herbert Stärker from Vienna (Austria), contributed to compile photographic field guides for the orchids of Africa and in particular South Africa (Johnson, Bytebier & Stärker 2015). Over a period of two decades he made regular trips to Africa to photograph orchids, including D. ver-rucosa. After many unsuccessful attempts to find this species, his wife joined a search expedition and rediscovered the long-lost orchid near Mtunzini in August 2011. Additional searches were conducted, leading to the discovery of another two plants in close proximity to the first. Over time he revisited these plants in the hope of photographing their fully opened flowers. No additional plants or populations were found after that. Due the sensitive nature of the rediscovery of these rare plants, Stärker and his colleagues kept their rediscovery quiet until more plants and populations could be found.
Unaware of the rediscovery of this orchid, the Custodians of Rare and Endangered Wildflowers (CREW) in association with the South African Botanical Society and the South African National Biodiversity Institute (SANBI) contacted the University of Zululand (UniZul) to request help to find and monitor some of Zululand's Red List plant species, of which D. verrucosa is one. Due to the ethical considerations surrounding research on such rare species, we decided that no vegetative or reproductive material needed to be collected during the initial stages. Therefore, no collection permits were requested from Ezemvelo KZN Wildlife. With the help of many students, a variety of methods were employed during the initial searches for D. verrucosa plants and populations. We initially followed the standard procedure used by CREW, which is to visit known historical localities and to conduct species-specific searches. All searches were conducted during the flowering-fruiting season of D. verrucosa, which was estimated to range from early August to late September. In some years we extended our searches into the month of October in the hope of finding dried fruit capsules. We spent approximately 195 man-hours in searches at known historical sites, covering approximately 6.5 ha over a period of five flowering seasons. Failing to record any plants, we adopted a grid-search strategy, during which ten fieldworkers searched a one-hectare sized area along predetermined gridlines ten metres apart, scanning the forest floor vegetation within mature Northern Coastal Forest (national vegetation type FOz7) (Mucina & Rutherford 2006). We spent approximately 672 man-hours in grid-searching mature forest, covering approximately 22.4 ha over a period of four flowering seasons. Failing to find D. verrucosa, we stratified the forest into dune crests, dune slopes and dune valleys before repeating our grid-search technique within each stratified unit again. This approach helped us to conduct more consistent searches within the relatively homogenous vegetation structure of each stratified unit. This follow-up search (90 man-hours covering 3 ha) was also done later (October) in the flowering-fruiting season, in the hope that a shift in search timing may lead to successful finds. Despite a total of approximately 960 man-hours spent searching an estimated 31.9 ha of forest floors south and east of Mtunzini over five consecutive flowering seasons, we did not locate any plants of D. verrucosa. Some of the above methods were employed concurrently, between September 2009 and October 2013. A chance discovery of a single plant was made in September 2013 by R.E. Mostert,
News of our discovery prompted Stärker to reveal the locality of their discovery to pool knowledge and manpower for more effective research and conservation efforts. At that time, these two localities (northeastern locality and southwestern locality) presented us with the opportunity to potentially monitor the four plants recorded. Three plants were recorded again the following year (2014), one in the northeastern locality and two in the southwestern locality. Whether these were the same plants as those recorded the previous years are unknown. No plants were recorded in the subsequent two years (2015 and 2016) at these two localities, despite very careful marking and thorough follow-up searches.
Objective
It was at this stage that we decided to change our strategy and follow Van Steenis' initial approach that contributed to chance discovery of D. verrucosa by focussing on ecosystem characteristics rather than the actual plants themselves. The aim of this study was therefore to produce a profile of indicators for the accurate identification of suitable habitat by analysing site characteristics of the two most recent discoveries. Such a profile could then be used to direct future search efforts better, or as a surrogate in the absence of actual field records when compiling conservation strategies and identifying target areas for the conservation of D. ver-rucosa. Whether these two localities truly represent optimal habitat or merely marginally suitable habitat is unknown at this stage.
Methods
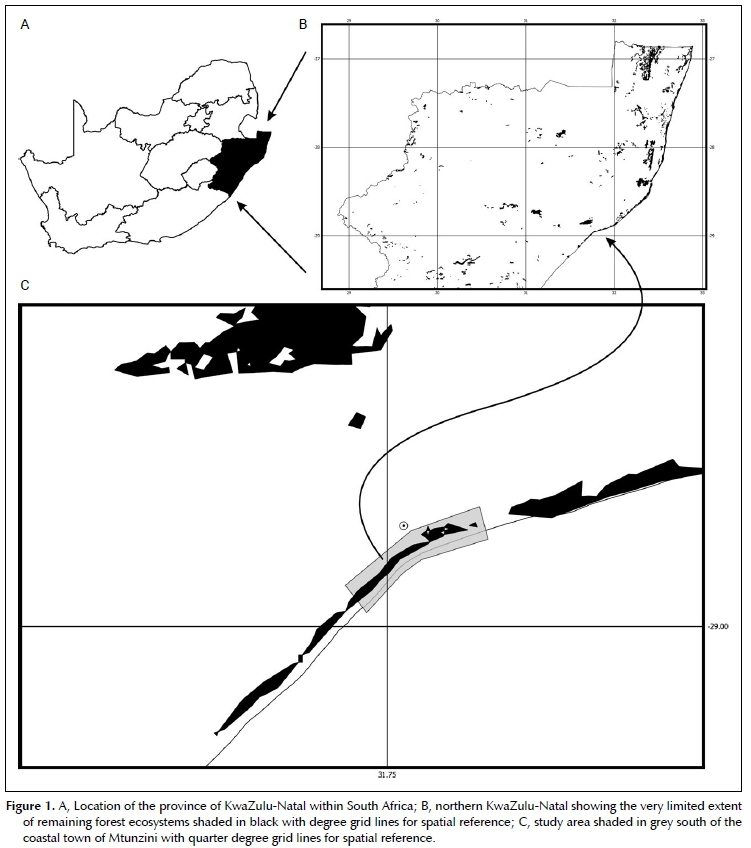
The habitat descriptions were compiled from both flo-ristic and environmental data gathered at the two most recent known localities of D. verrucosa. The two localities are approximately 2.3 km apart from one another. Field surveys were conducted while the plants were flowering/fruiting (2011-2012), before their subsequent disappearance. Due to the very sensitive nature of locality information of rare and endangered species, the exact localities are not provided here, with only an approximate locality of the study area presented in Figure 1 (straddling quarter degree squares 2831DC and 2831DD) (Larsen et al 2009). Exact localities were provided to the provincial conservation authority, Ezemvelo KZN Wildlife. For the purposes of this paper, we differentiate between the two localities by simply referring to them as the northeastern locality and south-western locality. It was decided to exclude historically known localities (1975 to 1993) from this habitat description due to the dynamic changes often associated with the forest floor conditions of Northern Coastal Forest (Lubke, Avis & Moll 1996; Zungu, Mostert & Mostert 2018). Vegetation structure was described in terms of the height and density of the various strata within the forest. Stratum classes were assigned according to the dominant height class structure system devised by Edwards (1983) for South African forest vegetation structure classification: upper canopy layer 15-25 m, short tree layer 8-15 m, tall shrub layer 3-8 m, short shrub layer 1-3 m, herbaceous layer 0.1-1.2 m.
The floristic composition of each stratum was described using the Braun-Blanquet approach. The Braun-Blanquet sampling method (Werger & Coetzee 1978) was specifically chosen for its international recognition as the most appropriate technique for the description of vegetation when based on total floristic composition (Brown et al. 2013). By using an internationally accepted standard, the data and vegetation description will be compatible and comparable with data from other regions and landscapes. Such comparability and compatibility are crucial for the regional and international coordination of vegetation and ecosystem conservation. Based on the recommendations by Brown et al. (2013), plot sizes were set at 400 m2 and were marked out in the field to ensure consistency. In all sample plots, each plant species was recorded and the cover-abundance value of each species determined using the modified Braun-Blanquet cover-abundance scale: r (very rare, with a negligible cover), + (present but not abundant, with a cover value of < 1% of the quadrat), 1 (numerous but covering less than 1% of the sample area), 2a (covering 5%-12% of the sample area), 2b (covering 13%-25% of the sample area), 3 (covering 25%-50% of the sample area), 4 (covering 50%-75% of the sample area), 5 (covering 75%-100% of the sample area) (Brown et al. 2013).
A wide range of environmental parameters were recorded and compared at the two localities. Distance from the average high-tide level along the nearby beach, as a measure of exposure to salt-clipping and wind-shearing from onshore wind and airborne sand particles, was calculated from georeferenced aerial imagery. Position and orientation descriptions within the vegetated dune cordon took into account whether a locality was situated on a dune crest, slope or valley and the number of dunes between it and the prograding beach. Soil texture and type was described using the standardised South African soil classification system (Fey 2010). Soil profile colour was described using the Munsell soil colour chart (Brady & Weil 2004). Thickness of the leaf-litter layer was measured in 10 mm increments. Texture of the leaf-litter was determined by measuring particle size (n = 100) at 10 mm above the soil surface. Composition of the leaf-litter on the forest floor was determined by a morphological analysis of decaying leaf fragments based on venation and trichome patterns. For this, 500 cm3 of organic material was collected from the upper layer leaf-litter layer between one and two metres away from the recorded D. verrucosa plants, taking care not to disturb the plants. Light penetration at forest floor level was determined by measuring the percentage of area covered in shadow and direct sunlight at 12:00 within the 400 m2
Braun-Blanquet vegetation quadrat. Hydrology was described based on the soil profile characteristics and the water holding capacity of the A horizon using the soil-sausage technique (Fey 2010). Slope was determined using a clinometer, while aspect was determined using a compass. Levels of ecosystem disturbance and fragmentation were described based on the degree of invasion by invasive alien plant species, the presence and extent of human activity, extent of animal impact on above-ground vegetation and leaf-litter movement, extent of damage to the herbaceous vegetation along the forest floor by falling trees or branches and the extent of damage to vegetation and leaf-litter by wind and water. The potential of localities to withstand storm damage were described based on a visual assessment of wind damage to plants and the redistribution of dead organic matter along the forest floor.
In addition to recording the above-mentioned environmental drivers of suitable habitat, some of the potential threats to the survival of D. verrucosa within one kilometre of the two localities were investigated and described. This included formal protection status, illegal logging, slash-and-burn agricultural practices, changes to natural burning regimes, human settlement, livestock grazing and browsing, impact of tourism, mining activities, invasive alien plant species, natural resource utilisation and management and illegal collection by traditional healers, nurseries and amateur collectors. Due to the low number of localities investigated (n = 2), no statistical correlation analyses were conducted on the data.
Results
Macro habitat description
The study area is located within the southernmost extremity of the Maputaland Centre of Plant Endemism (Van Wyk & Smith 2001) and within the Maputaland-Pondoland-Albany Hotspot, which is listed as one of the world's 36 most important biological hotspots (Myers et al. 2000). These hotspots are areas containing more than 1 500 endemic vascular plant species with less than 30% of their original vegetation still intact. The Maputaland-Pondoland-Albany Hotspot in particular contains approximately 1 900 endemic plant species, with less than 25% of its original vegetation still intact by 2004 due to anthropogenic activities (Steenkamp et al. 2004). The rate of ecosystem transformation has since increased (CEPF 2010).
Both sample localities fall within the Northern Coastal Forest vegetation type (FOz7) of the Indian Ocean Coastal Belt Biome (Mucina & Rutherford 2006). These forests are associated with wind-blown sand dunes that form some of the youngest geological formations in South Africa (<10 000 years old) (Gaugris et al. 2004). Within the study area, this specific dune cordon is currently expanding in the direction of the ocean at a rate of approximately one new dune per decade (Van Daalen et al. 1986; Weisser, Garland & Drews 1982; Weisser & Muller 1983) and is stabilised by one of the few examples of primary succession of vegetation in South Africa (Todd 1994; Zungu et al. 2018).
The overall climate in the study area is subtropical with hot, humid summers and cool moist winters (Boyes et al. 2011; Nevill & Nevill 1995; Rawlins 1991). The mean minimum and maximum temperatures are 11°C (July) and 30°C (December) respectively. Rainfall peaks twice a year with most received in summer (60% in November-March) and the rest in winter (40% in June-July) with a mean annual total of 1 100 mm. (Rawlins 1991; Tyson & Preston-White 2000). Relative humidity in summer usually exceeds 80% and during winter it often ranges from 50% to 60%. Potential evaporation exceeds average precipitation for all months except January, February and December and varies from 190 mm in January to 84 mm in June (Schulze 1982).
Microenvironment description
Abiotic parameters
The northeastern and southwestern localities both lie between the third and fourth dune series, respectively 370 m and 460 m from the average high-tide mark on the beach. Both are associated with relatively flat to slightly concave inter-dune valley topography of the dune cordon system. These protected valleys accumulate very large quantities of leaf-litter, leading to a very thick and stable carpet (40-80 mm) of decaying organic material on the forest floor (Moll 1972; Van Aarde, Smit & Claassens 1998). The leaf-litter is highly decomposed with a high density of fungal hyphae binding organic material into a mat. Although organic particle sizes range from microscopic dust to decaying tree trunks, the general texture of this organic carpet (at 10 mm above the soil surface) is relatively fine (1-7 mm in diameter). The morphological analysis of decaying leaf fragments (based on venation and trichome patterns) indicated that most leaf-litter comes from Isoglossa woodii, which often forms monotypic stands within the forest understorey where D. verrucosa grows.
Due to the relatively high elevation (15-25 m above sea level) of these specific dune valleys and the deep sandy soils associated with D. verrucosa, water drainage is very fast with no accumulation of free surface water, even during very heavy rainfall events (Watkeys, Mason & Goodman 1993). This results in relatively dry, aerobic conditions within the leaf-litter layer above the soil (Van Aarde et al. 1998). The soil associated with both sites is classified as cover sands (Fey 2010), with a mean grain size of 330 μm (Ware 2001) and highly leached (Watkeys et al. 1993). The upper 15 mm of sand directly below the leaf litter is greatly enriched with finely decomposed organic particles, resulting in colouration described by the Munsell Soil Colour Chart as 10YR3/3 dark yellow brown (Munsell 1980). Sand colour at a depth of 15-450 mm can be described as 10YR6/2 light brownish grey, while sand from 450-1 200 mm matched 10YR8/6 yellow of the Munsell chart.
The structural complexity of these forests and their topographic position result in a high level of protection from wind exposure, with no evidence of wind damage recorded within the forest understorey of the sampling quadrats. The only disturbance recorded within the forest floor of the sample quadrats were tracks, communal defecation middens and browsing damage to plants from red duiker (Cephalopus natalensis) and bushbuck (Tragelaphus sylvaticus). However, severe disturbances caused by indigenous bush pigs (Potamochoerus larva-tus) and a small group of feral pigs (Sus scrofa) within the forest floor were recorded in nearby forest patches.
Vegetation structure
Both localities fall within mature Northern Coastal Forest vegetation with multiple strata of interlocking canopies resulting in a total canopy cover of 100%. The fourth dune generally contains some of the oldest and structurally most complex forest vegetation within this dune cordon (Von Maltitz, Van Wyk & Everard 1996; Zungu et al. 2018). Beyond the fourth dune, most forest vegetation has been transformed by anthropogenic activities, with only very small sections of the fifth and sixth dunes still covered by the original coastal forest. The upper canopy on the fourth dune is approximately 20 m high and contributes to 40% of the total vegetation cover. This upper canopy interlocks with a shorter tree layer of approximately 8-15 m tall with a 60% cover. The tall shrub layer (3-8 m tall) is relatively sparse with a sub-canopy cover of approximately 20%. A very large number of lianas enter the canopy and an estimation of their contribution to the total canopy cover was not attempted. The herbaceous layer (0.1-1.2 m tall) is relatively dense (40-80% sub-canopy cover) and very dynamic. The dominant species within this layer is the long-lived, synchronously monocarpic herbaceous Isoglossa woodii, which grows for approximately 4-7 years before mass-flowering, setting seed and dying (Griffiths et al. 2010). This results in extreme structural changes within this vegetation layer at the end of each synchronised flowering cycle.
Floristic composition of the associated plant community
The uppermost stratum contains many old and reproductively mature trees (15-20 m high, 40% canopy cover) and is dominated by species such as Mimusops caffra, Sideroxylon inerme, Albizia adianthifolia, Ficus natalensis and Ekebergia capensis. Other prominent species within this layer include Cussonia spicata, Vepris lanceolata, Harpephyllum caffrum and Trichilia emetica. Zungu et al. (2018) described similar vegetation within the Umlalazi Nature Reserve as the Carissa bispinosa-Mimusops caffra climax coastal dune forest community.
The next stratum of shorter trees (8-15 m, 60% canopy cover) is dominated by species such as Brachy-laena discolor, Psydrax obovata, Dovyalis longispina, Gymnosporia arenicola, Protorhus longifolia, Tricalysia sonderiana, Apodytes dimidiata, Deinbollia oblongifolia and Clerodendrum glabrum. Other prominent species within this layer include Maytenus acuminata, Teclea gerrardii, Catunaregam obovata, Garcinia gerrardii, Maytenus procumbens, Tabernaemontana ventricosa, Chaetachme aristata, Olea woodiana, Bridelia micrantha, Putterlickia verrucosa, Grewia caffra and Canthium inerme.
Below the above stratum, a sparse layer of tall shrubs and low trees (3-8 m, 20% canopy cover) are dominated by Carissa bispinosa, Monanthotaxis caffra, Ochna serrulata, Peddiea africana, Eugenia capensis, Euclea natalensis, Cussonia zuluensis, Pavetta revoluta, Scutia myrtina, Allophylus natalensis, Kraussia floribunda and Dracaena aletriformis.
The lowest stratum is dominated by the herbaceous species Isoglossa woodii. This long-lived, synchronously monocarpic species ranges from 0.1 m tall during its first year of life, to 1.2 m tall by the time it flowers in mass approximately 4-7 years later. At the peak of its growth cycle, it covers between 40 and 80% of the forest floor, while the synchronised mass die-off event at the end of the growth cycle renders the forest floor almost devoid of live plant cover. Other species were recorded only at very low densities and included the ground orchid Cheirostylis gymnochiloides, the fern Microsorum scolopendria, the grass Oplismenus hirtellus, and other herbaceous forbs such as Behnia reticulata and Aneilema aequinoctiale.
Numerous woody lianas and herbaceous climbers penetrate various strata within the forest and include species such as Rhoicissus rhomboidea, Rhoicissus tomentosa, Rhoicissus sessilifolia, Dalbergia armata, Smilax anceps, Landolphia capensis, Dioscorea cotinifolia, Asparagus falcatus, Adenia gummifera, Cissampelos torulosa, Senecio tamoides, Senesio deltoideus and Secamone filiformis. A wide variety of epiphytes were recorded and include the fern Microsorum punctatum, the orchids Polystachya sandersonii, Cyrtorchis praetermissa, Cyrtorchis arcuata and Ansellia africana, as well as the parasitic mistletoe Erianthemum dregei.
Some of the invasive alien species recorded within the sample plots included Lantana camara, Chromolaena odorata and Rivina humilis. Although these species were only recorded at very low densities, large sections of adjacent forest patches have been completely invaded by Chromolaena odorata and Rivina humilis, transforming the forest floor conditions.
Discussion
As is the case with most taxonomic and phytosociological research, this habitat study should be regarded as a descriptive study. Due to the paucity of D. verrucosa localities (n = 2), no meaningful statistical correlation analyses could be conducted on the data. Whether the two sample localities represent distinct populations or only one is not known. Unfortunately, this scenario is all too common for many of South Africa's rare and endangered species (SANBI 2010). Suppressing the publication of such descriptive studies due to their lack of statistical verification will result in valuable information never being incorporated into conservation strategies and management plans (Pierce et al. 2005).
Critical environmental factors shaping suitable habitat for D. verrucosa are those that affect conditions along the forest floor. Based on the limited ecological information provided by Stewart and Hennessy (1980) and the habitat description presented in this study, we suggest that the following be considered to be critical parameters for suitable D. verrucosa habitat:
• Old-growth Northern Coastal Forest in pristine condition. Along this pro-grading shoreline, such forest is restricted to between the third to fourth dune from the beach, but along a degrading beach these old-growth forest patches may occur closer to the beach.
• High relative humidity under a dense multi-stratum canopy (100%) that buffers daily fluctuations in extreme temperature, light exposure, wind speed and desiccation.
• The co-occurrence of Isoglossa woodii (or similar perennial herbaceous species) as the dominant species in order to create medium-term (4-7 years) stable conditions within the lowest stratum of the forest vegetation.
• A thick carpet of fine textured organic matter mostly comprising of soft-leafed herbaceous material, such as that provided by Isoglossa woodii.
• Topography that results in the net accumulation of organic material with in situ decomposition.
• Low levels of disturbance within the leaf-litter mat and protection from damage by wind, water, large animals and invasive alien plant species.
• Fast draining sandy soil to ensure aerobic conditions, with no free-standing water along the forest floor. Soil texture, structure and nutrition seem to be less important to D. verrucosa since the roots and tubers are situated within the leaf-litter and were not recorded to enter the soil itself.
• High density of fungal hyphae in the soil which are parasitised by D. verrucosa.
As a by-product of the habitat analysis, a wide range of threats to the long-term survival of D verrucosa at these two localities, as well as at other similarly suitable habitats in the immediate surroundings, were recorded. These include:
• Lack of effective protection of remaining suitable habitat, with only one of the sites formally protected (Umlalazi Nature Reserve).
• Habitat degradation and fragmentation of the remaining Northern Coastal Forests due to unlawful logging, slash-and-burn agricultural practices, burning of adjacent rangelands to improve grazing for cattle, formal and informal settlements, recreational hiking trails and open-cast mining for heavy metals.
• Invasive alien plant species such as Chromolaena odorata (Ambika 2002; Hu & Zhang 2013) and Rivina humilis (Ravi et al. 2020), which are changing the forest floor characteristics dramatically due to competition and allelopathic effects of the secondary metabolites they contain.
• Increased chances of forest fires due to the forest margin changing from being fire-proof to fire-prone under the influence on Chromolaena odorata and Lantana camara.
• Current efforts to mechanically eradicate Chromolaena odorata and Rivina humilis in the immediate vicinity leads to the severe disruption of the leaf-litter on the forest floor.
• The impact of a small group of feral pigs.
• Close proximity to footpaths, making them vulnerable to accidental trampling and illegal collection.
• The negative effects of environmental stochasticity on small populations.
It is unknown whether the dramatic changes caused by the cyclical synchronised mass die-off of Isoglossa woodii after flowering present D. verrucosa with critical opportunities (e.g. easier cross pollination) or critical obstacles (e.g. exposure to unfavourable environmental conditions). Nonetheless, it is likely to have a profound effect on the flowering, pollination and seeding opportunities for D. verrucosa.
It is unknown whether D. verrucosa is a long-lived intermittent flowering species or whether they are simply short-lived reseeders. Autecological studies are needed to answer such questions. The precise environmental cues that trigger the tuber to produce a flowering stalk is still unknown. We have been trying for the last ten years to predict flowering times without any success. The answer may lie in the tubers it produces, enabling them to use previous years' resources for reproduction at a more flexible time frame to synchronise their phenology with environmental variables, such as the emergence of a suitable pollinator. The most detailed and comprehensive visual account of D. verrucosa is still the sketch made by E.F. Hennessy (Stewart & Hennessy 1981), based on observations made from a few plants she grew and maintained in a shoe box.
Although a small number of Hymenoptera and Diptera species have been recorded landing on the flowers during our surveys, no pollination behaviour was recorded. With flowers staying open for only a few hours before closing again, and the recorded spatial distribution of plants, successful crosspollination by an insect seems to be a truly herculean accomplishment. The heavy use of insecticides, herbicides and fertilisers within Zululand may have already had dire consequences for many potential pollinator species (Rother et al. 2008). Although the cleistogamous self-fertilisation recorded in D. verrucosa may provide a back-up chance to ensure a next generation, regular inbreeding as a result of continued failure to cross pollinate, will make the species less adaptable and more vulnerable to future environmental changes.
The combination of the many stochastic and deterministic events recorded in this study drive habitat change at rates beyond the ability of D. verrucosa to adapt its life history strategies for long term survival (Pressey et al. 2007). Managing anthropogenic degradation and fragmentation of suitable habitat should be our first priority when compiling conservation strategies of this very unique and rare species (Margules & Pressey 2000).
Conservation status revision
Raimondo et al. (2009) conducted a formal conservation status revision for D. verrucosa in 2009 and assigned the status of 'Vulnerable due to a population with a very restricted area of occupancy (less than 20 km2) and with locations (typically five or fewer) such that it is prone to the effects of human activities or stochastic events within a very short time period in an uncertain future, and is thus capable of becoming Critically Endangered or even Extinct in a very short time period (VU D2)'. After our own evaluation of the current situation using the formal IUCN criteria for assigning conservation status to species (IUCN 2020), we propose that this species be assigned the conservation status of Critically Endangered Category B2a and D. We base our recommendation on the following: IUCN Criterion B2a - Geographic area of occupancy estimated for D. verrucosa is less than 10 km2, and estimates indicate (a) severely fragmented population, as well as IUCN Criterion D - Population size estimated to number fewer than 50 mature individuals. Based on the difficulty to find and monitor these seemingly fleeting populations, we regard the population trend as unstable.
Suitable habitat for D. verrucosa is very limited within the region and under severe threat of degradation and fragmentation due to the long list of threats provided above. This is exacerbated by the limited effective protection of mature Northern Coastal Forests along this section of KwaZulu-Natal. At present, only the north-eastern locality falls within a formally protected area (Umlalazi Nature Reserve).
Conclusion
Traditional search techniques are inadequate for population monitoring programmes of rare and cryptic species such as D. verrucosa. This results in a lack of much needed quantitative data to better analyse and understand their ecology (Rouget et al. 2003). In the absence of such data, this study provides conservation agencies with a detailed description of its current known habitat, presenting a profile of indicators for the accurate identification of suitable habitat. Such information can now be used to direct future search efforts, or it can act as a surrogate in the absence of actual field records when compiling conservation strategies and management plans for D. verrucosa (Hannah et al. 2002; Pierce et al. 2005). This newly developed suitable-habitat-profile search strategy is currently being implemented by re-mapping all suitable habitat before searching for new D. verrucosa localities and colonies.
Acknowledgements
Herbert Stärker for sharing his knowledge and experiences regarding the rediscovery of D. verrucosa.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
THCM: Project coordinator, research design, interpretation of results, discussion and conclusion.
REM: Data collection, vegetation analysis and description, conservation status revision.
References
Ambika, S.R., 2002, 'Allelopathic Plants: Chromolaena odorata (L)', Allelopathy Journal 9(1): 35-41, [ Links ]
Botha, G.A., 1997, 'The Maputaland Group: a provisional lithostratigraphy for coastal KwaZulu-Natal', in Maputaland focus on the Quaternary evolution of the south-east African coastal plain, International Union for Quaternary Research Workshop Abstracts, Council for Geoscience, Private Bag X 112, Pretoria, South Africa, pp. 21-26. [ Links ]
Boyes, L.J., Gunton, R.M., Griffiths, M.E. & Lawes, M.J., 2011, 'Causes of arrested succession in coastal dune forest', Plant Ecology 212: 21-32, DOI: 10.1007/s11258-010-9798-6 [ Links ]
Brady, N.C. & Weil, R.R., 2004, Elements of the Nature and Properties of Soils. Prentice Hall, Upper Saddle River. [ Links ]
Brown, L.R., Du Preez, P.J., Bezuidenhout, H., Bredenkamp, G.J., Mostert, T.H.C. & Collins, N.B., 2013, 'Guidelines for phytosociological classifications and descriptions of vegetation in southern Africa', Koedoe 55(1), Art. #1103, 10 pages, http://dx.doi.org/10.4102/koedoe.v55i1.1103 [ Links ]
CEPF, 2010, 'Ecosystem Profile: Maputaland-Pondoland-Al-bany Biodiversity Hotspot', Critical Ecosystem Partnership Fund, occasional report 23 April 2010, pp. 135 [ Links ]
Edwards, D., 1983, 'A broad-scale structural classification of vegetation for practical purposes', Bothalia 14: 705-712. https://doi.org/10.4102/abc.v14i3/4.1231 [ Links ]
Fey, M.V., 2010, Soils of South Africa. Cambridge University Press, Cape Town. [ Links ]
Gaugris, J.Y, Matthews, W.S., Van Rooyen M.W. & Bothma. J. Du P., 2004, 'The vegetation of Tshanini Game Reserve and a comparison with equivalent units in the Tembe Elephant Park in Maputaland, South Africa', Koedoe 47(1): 9-29. Pretoria, https://doi.org/10.4102/koedoe.v47i1.67 [ Links ]
Griffiths, M.E., Tsvuura, Z., Franklin, D.C. & M.J. Lawes, M.J., 2010, 'Pollination ecology of Isoglossa woodii, a long-lived, synchronously monocarpic herb from coastal forests in South Africa', Plant Biology (Stuttg.) 12(3): 495-502. https://doi:10.1111/j.1438-8677.2009.00222.x [ Links ]
Hannah, L., Midgley, G.F., Lovejoy, T., Bond, W.J., Bush, M., Lovett, J.C., Scott, D. & Woodward, F.I., 2002, 'The conservation of biodiversity in changing climate', Conservation Biology 16(1): 264-268, DOI: 10.1046/J.1523-1739.2002.00465.x [ Links ]
Hu, G. & Zhang, Z., 2013, 'Allelopathic effects of Chromolae-na odorata on native and non-native invasive herbs', Journal of Food, Agriculture & Environment 11(1): 878-882. [ Links ]
IUCN 2020, 'The IUCN Red List of Threatened Species', Version 2020-1, https://www.iucnredlist.org. [ Links ]
Johnson, S.D., Bytebier, B. & Stärker, H., 2015, Orchids of South Africa. Struik Nature, Cape Town, South Africa. [ Links ]
Kalkman, K. 1990, 'Van Steenis remembered', in Baas, P., Kalkman, K. and Geesink, R. (eds) 1990 Proceedings of the Flora Malesiana Symposium: The plant diversity of Male-sia, Leiden University, Kluwer Academic Publishers, Dordrecht, The Netherlands August 1989, pp. 1-9. [ Links ]
Larsen, R., Holmern, T., Prager, S. D., Maliti, H. & R0skaft, E., 2009, 'Using the extended quarter degree grid cell system to unify mapping and sharing of biodiversity data', African Journal of Ecology, 47(3): 382-392, DOI: 10.1111/j.1365-2028.2008.00997.x [ Links ]
Lubke, R.A., Avis, A.M. & Moll. J.B., 1996, 'Post-mining rehabilitation of coastal sand dunes in Zululand, South Africa', Landscape and Urban Planning 34: 335-345. [ Links ]
Margules, C.R. & Pressey, R. L., 2000, 'Systematic conservation planning', Nature 405: 243-253. [ Links ]
Miura, C., Yamaguchi, K., Miyahara, R., Yamamoto, T., Fuji, M., Yagame, T., Imaizumi-Anraku, H., Yamato, M., Shigenobu, S. & Kaminaka, H., 2018, 'The mycoheterotrophic symbiosis between orchids and mycorrhizal fungi possesses: Major components shared with mutualistic plant-mycorrhizal symbioses', Molecular Plant-microbe Interactions 31(10):1032-1047, doi: 10.1094/MPMI-01-18-0029-R. [ Links ]
Moll, E. J., 1972, 'A Preliminary Account of the dune communities at Pennington Park, Mtunzini, Natal', Bothalia 10(4): 615-626 [ Links ]
Mucina, L. & Rutherford, M.C., (eds) 2006, 'The vegetation of South Africa, Lesotho and Swaziland', Strelitzia 19, South African National Biodiversity Institute, Pretoria. [ Links ]
Munsell Color Company, 1980, 'Munsell Book of Color', Munsell Color, Baltimore. [ Links ]
Myers, N., Mittermeier, R.A., Mittermeier, C.G., da Fonseca, G.A.B. & Kent, J., 2000, 'Biodiversity hotspots for conservation priorities', Nature 403: 853-858. [ Links ]
Neumann, F.H., Scott, L., Bousman C.B. & Van As, L., 2010, 'A Holocene sequence of vegetation change at Lake Eteza, coastal Kwazulu-Natal, South Africa', Review of Palaeobotany and Palynology 162: 39-53. [ Links ]
Nevill, H. & Nevill E.M., 1995, 'A survey of the Culicoides (Diptera: Ceratopogonidae) of the UMlalazi Nature Reserve in Zululand, South Africa, with notes on two species biting man', Onderstepoort Journal of Veterinary Research 62: 51-58. [ Links ]
Pierce, S.M., Cowling, R.M. Knight, A.T.T., Lombard, A.T., Rouget, M. & Wolf, T., 2005, 'Systematic conservation planning products for land-use planning: Interpretation for implementation', Biological Conservation 125: 441-458. [ Links ]
Pressey, R.L., Cabeza, M., Watts, M.E., Cowling, R.M. & Wilson, K.A., 2007, 'Conservation planning in a changing world', Trends in Ecology and Evolution 22(11): 584-592, doi: 10.1016/j.tree.2007.10.001 [ Links ]
Raimondo, D., Von Staden, L., Foden, W., Victor, J.E., Helme, N.A., Turner, R.C., Kamundi, D.A. & Manyama, P.A., 2009, 'Red List of South African Plants', Strelitzia 25, South African National Biodiversity Institute, Pretoria. [ Links ]
Ravi, R.K., Anusree, S., Balachandar, M. & Thangavelu, M., 2020, 'Plastic responses that confer invasiveness to Rivina humilis L.', Acta Ecologica Sinica, DOI: 10.1016/j.chnaes.2020.01.001 [ Links ]
Rawlins, B.K., 1991, A geohydrological assessment of the behaviour and response of the Zululand coastal plain to both environmental influences and human activity, MSc dissertation, University of Zululand. [ Links ]
Rother, H., Hall, R. & London, L., 2008, 'Pesticide use among emerging farmers in South Africa: contributing factors and stakeholder perspectives', Development Southern Africa 25(4): 399-424. [ Links ]
Rouget, M., Richardson, D.M., Cowling, R.M., Lloyd, J.W. & Lombard, A.T., 2003, 'Current patterns of habitat transformation and future threats to biodiversity in terrestrial ecosystems of the Cape Floristic Region, South Africa', Biological Conservation 112(1): 63-85. [ Links ]
SANBI 2010, Red List of South African plants, South African Biodiversity Institute, Pretoria, http://redlist.sanbi.org/ [ Links ]
Schulze, R.E., 1982, 'Agrohydrology and agroclimatology of Natal', Agricultural Catchments Research Unit Report 14, University of Natal, Pietermaritzburg. [ Links ]
Soil Classification Working Group, 1991, 'Soil classification - A taxonomic system for South Africa', Memoirs of the Agricultural Resources South Africa 15, Department of Agricultural Development, Pretoria. [ Links ]
Steenkamp, Y., Van Wyk, B., Victor, J., Hoare, D., Smith, G., Dold, T., & Cowling, R., 2004, 'Maputaland-Pondoland-Albany'. In: R.A. Mittermeier, P. Robles Gil, M. Hoffmann, J.D. Pilgrim, T.M. Brooks, C.G. Mittermeier, & G.A.B. da Fonseca (eds.), Hotspots Revisited: Earth's Biologically Richest and Most Endangered Ecoregions, pp. 219-228. Monterrey, Mexico: CEMEX [ Links ]
Stewart, J. & Hennessy, E.F., 1980, 'Didymoplexis verrucosa - A new saprophytic orchid from South Africa', American Orchid Society Bulletin 49(8): 836-841. [ Links ]
Stewart, J. & Hennessy, E.F., 1981, Orchids of Africa: A Select Review, Houghton Mifflin Publishers, University of Michigan, pp 159. [ Links ]
Todd, C.B., 1994, A comparison of the reproductive strategies of key species of a prograding dune system in the Mlalazi Nature Reserve, Natal, MSc dissertation, Rhodes University. [ Links ]
Tyson, F.D. & Preston-Whyte, R.A., 2000, 'The weather and climate of southern Africa', Oxford University Press, Cape Town. [ Links ]
Van Aarde, R.J., Smit, A.M. & Claassens, A.S., 1998, 'Soil characteristics of rehabilitating and unmined coastal dunes at Richards Bay, KwaZulu-Natal, South Africa', Restoration Ecology 6(1): 102-110. [ Links ]
Van Daalen, J.C., Geldenhuys, C.J., Frost, P.G.H. & Moll, E.J., 1986, 'A rapid survey of forest succession at Mlalazi Nature Reserve', Foundation for Research Development CSIR, Occasional Report 11. [ Links ]
Van Wyk, A.E. & Smith, G.F., 2001, 'Regions of floristic endemism in southern Africa', Umdaus Press, Pretoria. [ Links ]
Victor, J.E., McMurtry, D., Grobler, L. & Burns, S. 2005, Didymoplexis verrucosa J.Stewart & Hennessy. National Assessment: Red List of South African Plants version 2020.1. Accessed on 2020/06/06. [ Links ]
Von Maltitz, G.P, Van Wyk, G.F. & Everard, D.A., 1996, 'Successional pathways in disturbed coastal dune forest on the coastal dunes in north-east Kwazulu-Natal, South Africa', South African Journal of Botany 62(4): 188-195. [ Links ]
Ware, C.I., 2001, Evolution of the northern KwaZulu-Natal coastal dune cordon; evidence from the fine-grained sediment fraction, MSc dissertation, University of Natal, South Africa. [ Links ]
Watkeys, M. K., Mason, Z. R. & Goodman, P S., 1993, 'The role of geology in the development of Maputaland, South Africa', Journal of African Earth Sciences 16(1): 205-221. [ Links ]
Weisser, P.J & Muller, R., 1983, 'Dune vegetation dynamics from 1937 to 1976 in the Mlalazi-Richards Bay area of Natal, South Africa', Bothalia 14(3): 661-667. [ Links ]
Weisser, PJ., Garland, I.F & Drews, B.K., 1982, 'Dune advancement 1937-1977 at the Mlalazi Nature Reserve, Mtunzini, Natal, South Africa, and a preliminary vegetation-succession Chronology', Bothalia, 14(1): 127-130. [ Links ]
Werger, M.J.A. & Coetzee, B.J., 1978, 'The Sudano-Zam-besian Region', in M.J.A. Werger (ed.), Biogeography and ecology in southern Africa, pp. 231-299, Junk, The Hague. [ Links ]
Zungu, N.S., Mostert, T.H.C. & Mostert, R.E., 2018, 'Plant communities of the uMlalazi Nature Reserve and their contribution to conservation in KwaZulu-Natal', Koedoe 60(1), a1449. https://doi.org/10.4102/koedoe.v60i1.1449 [ Links ]
 Correspondence:
Correspondence:
Dr T.H.C. Mostert
E-mail: MostertT@unizulu.ac.za
Submitted: 9 June 2020
Accepted: 17 February 2021
Published: 8 July 2021


{kind=link}