










Serviços Personalizados
Artigo
 Inglês (pdf)
Inglês (pdf)
 Artigo em XML
Artigo em XML Referências do artigo
Referências do artigo
Indicadores

Links relacionados
-
 Citado por Google
Citado por Google -
 Similares em Google
Similares em Google
Compartilhar

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
versão On-line ISSN 2311-9284
versão impressa ISSN 0006-8241
Bothalia (Online) vol.51 no.1 Pretoria 2021
http://dx.doi.org/10.38201/btha.abc.v51.i1.11
SHORT COMMUNICATION
A floristic assessment of grassland diversity loss in South Africa
M. MullerI; S.J. SiebertI; B.R. NtlokoII; F. SiebertI
IUnit for Environmental Sciences and Management, North-West University, Private Bag X6001, Potchefstroom 2520, South Africa
IILetseng Diamonds, cnr Kingsway and Old School Road Maseru, P.O. Box 12508, Maseru 100 lesotho
ABSTRACT
BACKGROUND: Land-use effects on grassland flora are difficult to predict due to poor understanding of species losses caused by transformation.
OBJECTIVES: To determine changes in species diversity and composition by comparing transformed with untransformed grassland.
METHODS: Floristics of paired plots were sampled within 18 transformed sites (representing agricultural and urban land-uses) and neighbouring untransformed grassland.
RESULTS: Endemic and threatened species were negatively affected by transformation, particularly species with belowground bud-banks and storage organs. Species composition, with clear shifts in dominant families, was changed by over 90% on average by transformation
CONCLUSION: Land-use transformation lead to the loss of native species and increased alien invasive species.
Introduction
Land-use change threatens the persistence of many grassland ecosystems worldwide (Bond 2016). Grasslands are hyper-diverse ancient ecosystems, habitats and communities, supporting many endemic and threatened species (Carbutt, Henwood & Gilfedder 2017). Habitat transformation threatens the integrity of these systems through soil disturbance and the removal of plant biomass and species, and the effect is widely recognised and measurable (Herben, Chytry & Klimesová 2016; Miller, Roxburgh & Shea 2011). The poor understanding of forb dynamics in grassland necessitates a closer look at flo-ristic change and whether land-use change leads to species losses or gains in transformed grassland (Veldman et al. 2015).
In South Africa the Grassland Biome covers approximately one third of the land surface (Carbutt et al. 2011). The extent of grassland is defined on the basis of vegetation structure, as well as environmental factors including mean summer rainfall and minimum winter temperatures (Mucina & Rutherford 2006). The Grassland Biome is one of the most at-risk South African biomes, with 40-60% irreversibly modified, and less than 3% formally protected (Little, Hockey & Jansen 2015). The intactness of unprotected South African grasslands is threatened as there is an increase in the intensity of agriculture and afforestation (O'Connor & Kuyler 2009; Botha et al. 2017) and urban and industrial development activities (Siebert, Van Wyk & Bredenkamp 2001; O'Connor & Kuyler 2009). Changes in composition, structure and functioning of these grasslands influence the ability to deliver fresh water, soil formation, climate regulation and reduction of disaster risk (Egoh et al. 2011), and in addition, probable loss of biodiversity and grassland production (Everson & Everson 2016).
O'Connor and Kuyler (2009) have meticulously investigated the impact of land-use on the biodiversity integrity of moist grasslands in South Africa and highlighted the loss of useful plants from an ecosystem services perspective. Our study focusses on biodiversity intact-ness in that it specifically considers loss of native floristic diversity. It places special emphasis on the indigenous forb component that is fast moving up the research agenda (Siebert & Dreber 2019).
Materials and methods
Study area
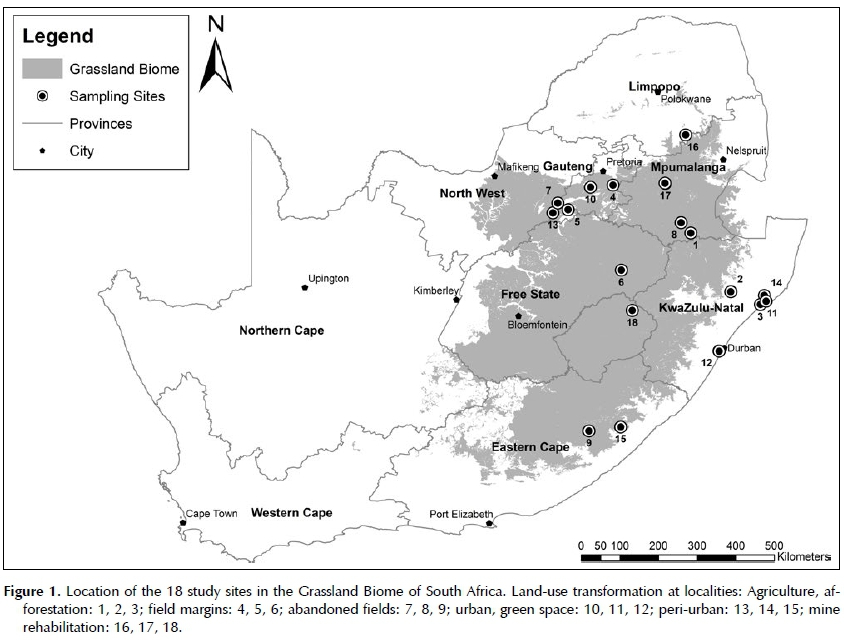
Eighteen study sites were selected in four bioregions of the Grassland Biome, as well as a tropical bioregion of the Indian Ocean Coastal Belt Biome of South Africa (Figure 1). The chosen grasslands occurred at altitudes ranging between 30 and 3 100 m above sea level, with ten sites between 1 000 and 1 800 m. The mean annual temperature for the grassland sites ranged from 10 to 21°C, with an overall mean of 16.3°C (median 15.9°C). June to August are the coldest months with mean frost days per annum varying between 0 and 96, with a mean of 25 (median 28) across all study sites (Mucina & Rutherford 2006). All sites experience summer rainfall ranging from 600 to 1 000 mm per year and a mean of 761 mm (median 717 mm) across sites. Twelve sites receive less than 800 mm per annum.
Field surveys
Two dominant land transformation types in the Grassland Biome were included in this study, namely agriculture and urbanisation (Neke & Du Plessis 2004). Floristic data were gathered from 18 sites. At each site, sampling was conducted in four plots in untransformed grassland, each paired with a plot in an adjacent transformed land-use (i.e. eight plots per site), no more than 150-250 m apart. All 144 plots were surveyed in late spring or early to mid-summer. Each 100 m2 plot was divided into 25 subplots of 4 m2 each to record species occurrence and abundance. Species were identified in the field and photos were taken for later confirmation. Floristic data from the subplots were combined to compile a total inventory for each 100 m2 plot.
Plant species nomenclature and classification follow Ranwashe (2019). Naturalised and invasive categories are according to Department of Environmental Affairs (2016). Life and growth forms of plant species were obtained from Germishuizen and Meyer (2003). Categories of threat were obtained from the Red Data List of South African Plants (South African National Biodiversity Institute 2017).
Species abundance (N) was calculated as the total number of individuals and species richness (S) as the total number of species within each 100 m2 plot. The Shannon-Wiener (H') and Pielou (J') indices were applied to plot data to calculate alpha diversity and evenness respectively. All above values were calculated using Primer (2007).
Data analysis
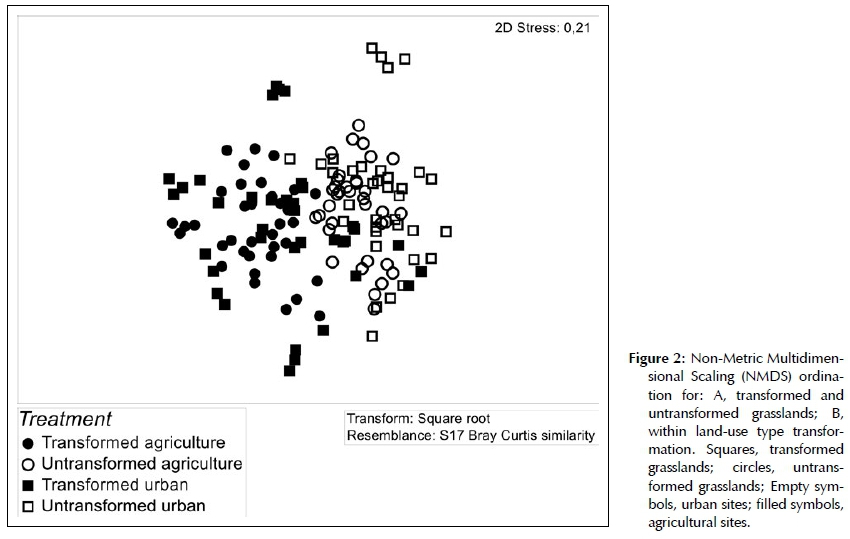
Non-Metric Multi-Dimensional Scaling (NMDS) analysis in Primer (2007) was used to explore changes in species composition between transformed and un-transformed grasslands. Permutational Multivariate Analysis of Variance (PERMANOVA) was performed using species abundance data. Analyses were conducted with 999 permutations using Bray-Curtis similarity and Type III sums of squares after a square root transformation of species data to reduce the influence of common species. To account for location variability in the paired, nested sampling design, plots were treated as a random variable nested within a transformation type (i.e. urban or agricultural), which were treated as the fixed factor. Pair-wise test results indicated the strength of the difference between transformed and untransformed plots.
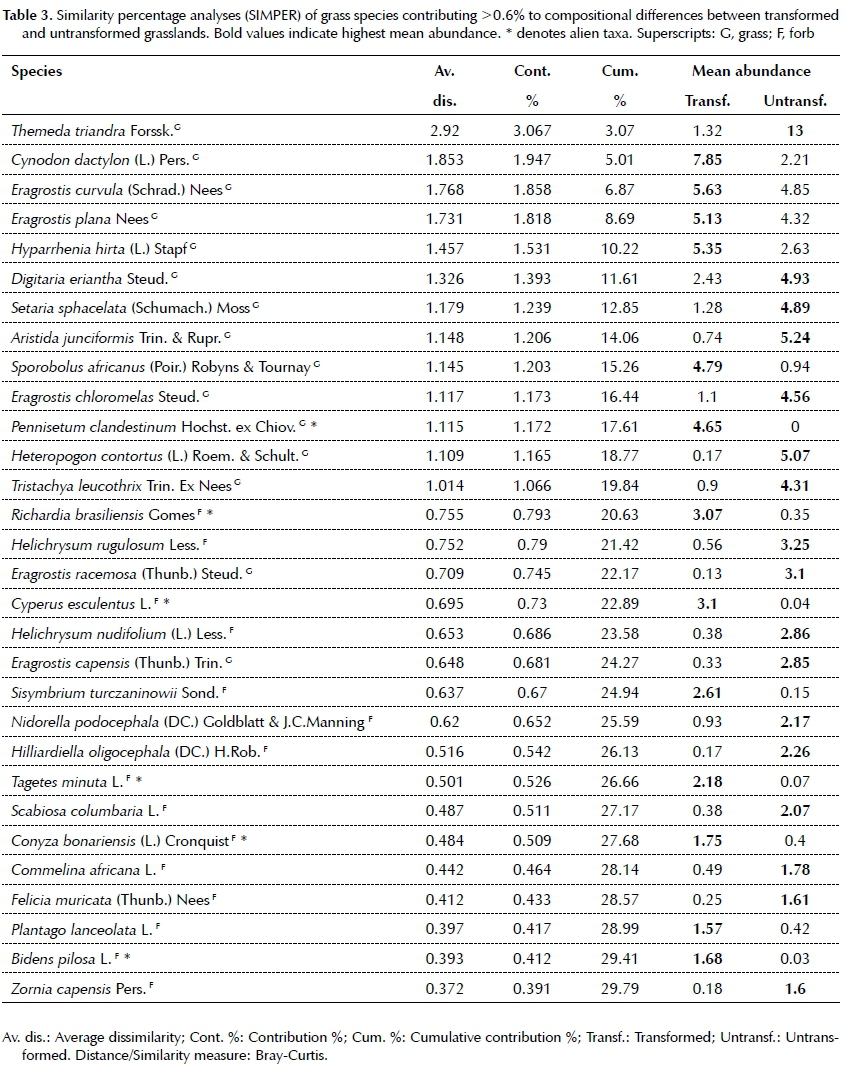
Similarity Percentage Analysis (SIMPER) was applied to determine which forb and grass species contributed the most to differences between transformed and obtained from Germishuizen and Meyer (2003). Categories of threat were obtained from the Red Data List of South African Plants (South African National Biodiversity Institute 2017).
Results
Floristics
Overall, 1 146 plant species were recorded, of which 144 were non-native. The untransformed grassland contained 962 species, which included 35 naturalised and 15 invasive taxa (5%), 175 South African endemics (18%) and 20 threatened species (2%). The transformed grasslands had 582 species, including 92 naturalised and 46 invasive taxa (24%), 47 South African endemics (8%) and six threatened species (1%).
The most prominent families in the localities were the Asteraceae, Poaceae, Fabaceae and Cyperaceae in order of most species diverse (Table 1), whereas the Poa-ceae were most abundant (Table 2). It is evident that transformation is less favourable to the Asteraceae and Fabaceae, and more beneficial to the Cyperaceae and especially the Poaceae.
Habitat transformation affects the number of species present per family. The geophytic Hyacinthaceae and Iridaceae showed the largest species losses (18 and 19 species respectively) when grassland is transformed and the weedy Amaranthaceae and Solanaceae benefitted in terms of species additions (9 and 5 species respectively; Table 1). Changes in the frequency of species is even more pronounced (Table 2). Five of the top ten families that have high frequencies of occurrence in un-transformed grassland become reduced in transformed grassland by 73%. These are replaced by five families, which in turn showed a 75% increase in transformed grassland (Table 2).
Composition
Changes in the species number and frequency of families is expected to have an effect on the composition of transformed grassland. The results from a NMDS revealed clustering that supports the untransformed and transformed grasslands as separate assemblages (Figure 2). Results from the pair-wise tests in PERMANOVA indicated a significant difference in floristic composition between transformed and untransformed grasslands in both urban (df = 70, t = 2.17, p = 0.001) and agricultural (df = 70, t = 2.88, p = 0.001) transformation types (Figure 2). Bray Curtis similarity measures in the PERMANOVA design reported a low 6.96% and 5.7% similarity in species composition between transformed and untransformed agricultural and urban grasslands respectively. This implies that transformation changed species composition in grasslands by ~90% on average. Fifteen most common grass species explained 21.26% of the dissimilarity between transformed and untrans-formed grasslands, with species such as Cynodon dacty-lon and Hyparrhenia hirta weighted towards the former and Digitaria eriantha and Themeda triandra towards the latter (Table 3). Comparatively the first 15 forbs species only contributed 8.52% to the dissimilarity, with species such as Cyperus esculentus and Richardia brasil-iensis weighted towards transformed, and Helichrysum rugulosum and Scabiosa columbaria towards untrans-formed grassland (Table 3).
Diversity
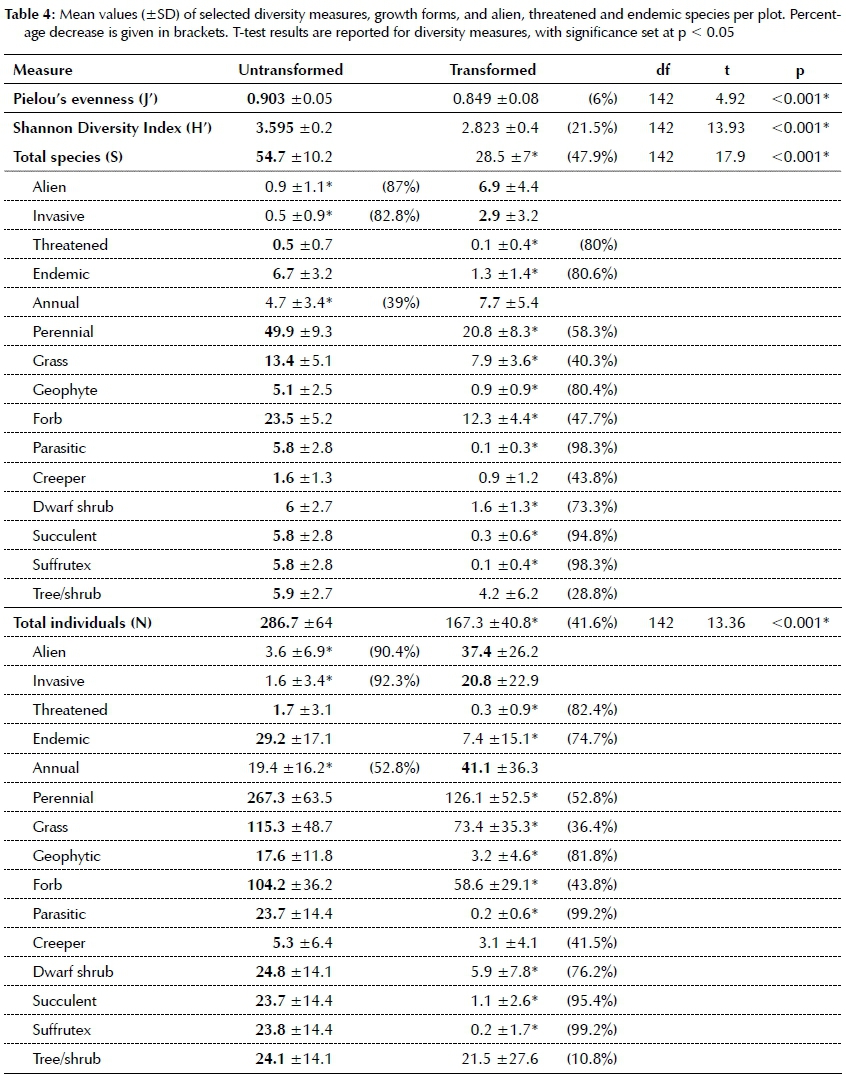
Changes in the species composition are expected to have an effect on species richness and diversity in transformed grassland. Simple paired t-tests revealed significantly lower diversity (for all measures, i.e. J', H', S and N) in the transformed grassland (p<0.001, Table 4). Species richness decreased by nearly 50%.
Status
The lower evenness in transformed grassland indicates uneven proportional contribution of individuals between species and is indicative of some species becoming more dominant and others becoming marginal. Dominance shifts can be ascribed to increased numbers of alien, invasive and annual species in the transformed grassland, with the former two being >80% lower in untransformed grassland (Table 4). Threatened and endemic species richness and abundance decreased by >80% in the transformed grassland (Table 4).
Growth forms
When aliens displace extant species (including endemics), certain growth forms become more or less prominent. Species losses (Table 4) were recorded for species with underground storage organs and bud-banks (USOs), such as geophytes (>80%), parasitic plants and suffrutices (>90%). Succulents were considered separately from the dwarf shrub growth form and were also decreased by over 90% in transformed grasslands (Table 4).
Discussion
Asteraceae and Fabaceae were the most dominant forb families in both transformed and untransformed grassland, a trend that has previously been reported (Botha et al. 2017). It can be deduced that regenerative traits, such as wind dispersal and seed dormancy for rapid colonisation (Asteraceae), ballochory and/or or endozo-ochory as seed dispersal traits, and resource acquisition traits, which increase resprouting capacity (Fabaceae), make them rather resilient to disturbance. However, the Amaranthaceae, Brassicaceae, Solanaceae and
Verbenaceae benefit from the transformation of grasslands and their numbers and dominance increase through the introduction of weedy, mostly alien, species. These groups are renowned for their ability to colonise frequently transformed man-made habitats (Pysek, Prach & Smilauer 1995). In contrast, certain families, such as the geophytic Hyacinthaceae and Iridaceae, were extensively disadvantaged by habitat transformation, as they are sensitive to soil disturbance because their bud-banks are belowground (Fidelis et al. 2014). The general trend is therefore one of species loss and displacement by a new flora, mostly annuals, with colonising traits better suited to a transformed environment, such as creepers, clonal plants and fruit or seed adapted for exozoochorous or anemochorous dispersal (Botha et al. 2017).
Many species of ancient grasslands are not tolerant to anthropogenic disturbance (Siebert 2011). Common native species disappeared completely where grasslands were transformed, such as the grasses Alloterop-sis semialata (R.Br.) Hitchc. and Schizachyrium san-guineum (Retz.) Alston, forbs Gerbera ambigua (Cass.) Sch.Bip. and Haplocarpha scaposa Harv., geophytes Ledebouria luteola Jessop and Hypoxis argentea Harv. ex Baker., dwarf shrubs Athrixia phylicoides DC. and Tephrosia capensis (Jacq.) Pers., and suffrutices Ele-phantorrhiza elephantina (Burch.) Skeels and Ziziphus zeyheriana Sond. Other species, such as the palatable and productive grass, Themeda triandra, which is considered a keystone species and indicator of undisturbed grassland (Snyman, Ingram & Kirkman 2013), were severely reduced. Many new, mostly alien species, enter the transformed system, providing species that can be considered indicators of disturbance (Morris & Scott-Shaw 2019), such as the prostrate and grazing-resistant, Richardia brasiliensis. This study confirmed the results of O'Connor (2015) that a substantial decrease in T. triandra and increase in R. brasiliensis are indicative of transformed grassland.
As previously shown, untransformed grasslands have greater plant species diversity (27%) and richness (92%) than transformed grasslands (O'Connor 2005; Siebert 2011). Species loss was specific to certain growth forms with underground organs specifically adapted to survive harsh winter conditions, drought and fire (Bond & Parr 2014; Bond 2016). The loss of these and other foundation species open up niches for colonisation by alien species (Prevéy et al. 2010). This is problematic, as loss of native species hampers the grassland ecosystem from fulfilling all its functions (Zavaleta et al. 2010). Overall, the situation in transformed grassland is not only one of species depletion, but increases in woody growth form dominance is predicted to become more pronounced in South African grasslands by 2030 (Gibson et al. 2018).
Conclusion
Major plant families remain floristically dominant after transformation, but there is a negative impact on overall phylogenetic diversity and the promotion of the Poa-ceae and Cyperaceae. Non-typical grassland families, with a wide array of disturbance-tolerant traits, show an increase in phylogenetic diversity, which is mainly a consequence of the introduction of alien weedy species.
Species composition of grassland was transformed by disturbance and is indicative of better adapted species entering the system or existing pre-adapted ones becoming more dominant due to competition release and/or altered microclimates and soils (Pysek, Prach & Smilauer 1995). This is evidenced by the proportion of grass species increasing, but a large reduction in forb species with USOs (Fidelis et al. 2014). No evidence was found for extensive woody encroachment in transformed areas.
This study set out to assess species loss due to transformation and, based on the current data set, it can be conclusively stated that grassland is severely impacted in terms of its species richness and diversity. These changes are of concern as grasslands have high economic value and support the wellbeing of humans by providing, among others, ecological infrastructure, carbon sinks, albedo surfaces, plant-based medicines, food plants and grazing for livestock (Bengtsson et al. 2019). Further studies are needed to determine whether these floristic shifts can still maintain and provide the ecosystem services that are expected from grasslands in South Africa.
Acknowledgements
Our appreciation to Dr Monique Botha, Dr Elandrie Davoren and Mr Paul Janse van Rensburg for making plot data available for this study, and to Mr Wynand Muller for producing the locality map. The South African National Biodiversity Institute, South African Environmental Observation Network, National Research Foundation of South Africa and Letseng Diamond Mine, Lesotho, provided financial support to the researchers and students involved in this project.
Authors' contributions
MM collected field data, conducted data analyses and contributed to the writing of the manuscript. SJS planned and coordinated the study, collected field data, conducted data analyses and co-wrote the manuscript. BRN planned and coordinated part of the study, collected field data, and conducted data analyses. FS collected field data and contributed to the writing of the manuscript.
Disclaimer
The views expressed in the submitted article are our own and not an official position of the institution or funder.
Source(s) of support
South African National Biodiversity Institute, South African Environmental Observation Network, National Research Foundation.
References
Bengtsson, J., Bullock, J.M., Egoh, B., Everson, C., Everson, T., O'Connor, T., O'Farrell, PJ., Smith, H.G. & Lindborg, R., 2019, 'Grasslands - more important for ecosystem services than you might think', Ecosphere 10(2), p.e02582. https://doi.org/10.1002/ecs2.2582 [ Links ]
Bond, W.J., 2016, 'Ancient grasslands at risk', Science 351 (6269),120-122. https://doi.org/10.1126/science.aad5132 [ Links ]
Bond, W.J., & Parr, C.L., 2010, 'Beyond the forest edge: ecology, diversity and conservation of the grassy biomes', Biological Conservation 143(10), 2395-2404. https://doi.org/10.1016/j.biocon.2009.12.012 [ Links ]
Botha, M., Siebert, S.J., Van den Berg, J., Ellis, S. & Dre-ber, N., 2017, 'Plant functional types differ between the grassland and savanna biomes along an agro-ecosystem disturbance gradient in South Africa', South African Journal of Botany 113, 308-317. https://doi.org/10.1016/j.sajb.2017.09.008 [ Links ]
Carbutt, C., Henwood, W.D. & Gilfedder, L.A., 2017, 'Global plight of native temperate grasslands: going, going, gone?', Biodiversity and Conservation 26(12), 2911-2932. https://doi.org/10.1007/s10531-017-1398-5 [ Links ]
Carbutt, C., Tau, M., Stephens, A. & Escott, B., 2011, 'The conservation status of temperate grasslands in southern Africa', The Grassland Society of Southern Africa 11(1), 1723. https://grassland.org.za/publications/grassroots/issues/february-2011/5%20201102%20Carbutt.pdf [ Links ]
Department of Environmental Affairs, 2016, National Environmental Management: Biodiversity Act 2004 (Act No. 10 of 2004) Alien and invasive species lists, Government Gazette 40166(864), Pretoria, pp. 31-104. [ Links ]
Everson, C.S. & Everson, T., 2016, 'The long-term effects of fire regime on primary production of montane grasslands in South Africa', African Journal of Range & Forage Science 33(1), 33-41. https://doi.org/10.2989/10220119.2015.1124922 [ Links ]
Fidelis, A., Appezzato-da-Glória, B., Pillar, V.D. & Pfadenhauer, J., 2014, 'Does disturbance affect bud bank size and be-lowground structures diversity in Brazilian subtropical grasslands?', Flora 209(2), 110-116. https://doi.Org/10.1016/j.flora.2013.12.003 [ Links ]
Germishuizen, G. & Meyer, N.L., 2003, Plants of southern Africa: an annotated checklist, National Botanical Institute, Pretoria. [ Links ]
Gibson, L., Münch, Z., Palmer, A. & Mantel, S., 2018, 'Future land cover change scenarios in South African grasslands - implications of altered biophysical drivers on land management', Heliyon 4(7), e00693. https://doi.org/10.1016/j.heliyon.2018.e00693 [ Links ]
Herben, T., Chytry, M. & Klimesová, J., 2016, 'A quest for species level indicator values for disturbance', Journal of Vegetation Science 27(3), 628-636. https://doi.org/10.1111/jvs.12384 [ Links ]
Little, I.T., Hockey, PA.R. & Jansen, R., 2015, 'Impacts of fire and grazing management on South Africa's moist highland grasslands: A case study of the Steenkampsberg Plateau, Mpumalanga, South Africa', Bothalia 45(1), Art. #1786. https://dx.doi.org/10.4102/abc.v45i1.178 [ Links ]
Miller, A.D., Roxburgh, S.H. & Shea, K., 2011, 'How frequency and intensity shape diversity-disturbance relationships', Proceedings of the National Academy of Sciences 108(14), 5643-5648. https://doi.org/10.1073/pnas.1018594108 [ Links ]
Morris, C.D. & Scott-Shaw, R., 2019, 'Potential grazing indicator forbs for two mesic grasslands in South Africa', Ecological Indicators, 107, 105611. https://doi.org/10.1016/j.ecolind.2019.105611 [ Links ]
Mucina, L. & Rutherford, M.C., 2006, The vegetation of South Africa, Lesotho and Swaziland, South African National Biodiversity Institute, Pretoria. [ Links ]
Neke, K.S. & du Plessis, M.A., 2004, 'The threat of transformation: quantifying the vulnerability of grasslands in South Africa', Conservation Biology 18, 466-477. https://doi.org/10.1111/j.1523-1739.2004.00157.x [ Links ]
O'Connor, T.G., 2005, 'Influence of land use on plant community composition and diversity in Highland Sourveld grassland in the southern Drakensberg, South Africa', Journal of Applied Ecology 42(5), 975-988. https://doi.org/10.111/j.1365-2664.2005.01065.x [ Links ]
O'Connor, T.G. & Kuyler, P., 2009, 'Impact of land use on the biodiversity integrity of the moist sub-biome of the grassland biome, South Africa', Journal of Environmental Management 90(1), 384-395. https://doi.org/10.1016/j.jenvman.2007.10.012 [ Links ]
Prevéy, J.S., Germino, M.J., Huntly, N.J. & Inouye, R.S., 2010, 'Exotic plants increase and native plants decrease with loss of foundation species in sagebrush steppe', Plant Ecology 207(1), 39-51. https://doi.org/10.1007/s11258-009-9652-x [ Links ]
Primer, 2007, Primer, Version 6, Primer-E Ltd, United Kingdom. [ Links ]
Pysek, P., Prach, K. & Smilauer, P., 1995, 'Relating invasion success to plant traits: an analysis of the Czech alien flora', in P Pysek, K. Prach, M. Rejmánek & M. Wade (eds), Plant invasions: General aspects and special problems, pp. 39-60, Academic Publishing, Amsterdam. [ Links ]
Ranwashe, F., 2019, BODATSA: Botanical Collections, v1.4, South African National Biodiversity Institute, viewed 15 May 2019, from http://ipt.sanbi.org.za/iptsanbi/re-source?r=brahms_online&v=1.4 [ Links ]
Siebert, F. & Dreber, N., 2019, 'Forb ecology research in dry African savannas: Knowledge, gaps, and future perspectives', Ecology & Evolution 9(13), 7375-7891. https://doi.org/10.1002/ece3.5307 [ Links ]
Siebert, S.J., 2011, 'Patterns of plant species richness of temperate and tropical grassland in South Africa', Plant Ecology and Evolution 144(3), 249-254. https://doi.org/10.5091/plecevo.2011.501 [ Links ]
Siebert, S.J., Van Wyk, A.E. & Bredenkamp, G.J., 2001, 'Ende-mism in the flora of ultramafic areas of Sekhukhuneland, South Africa', South African Journal of Science 97(11): 529-532. https://journals.co.za/content/sajsci/97/11-12/EJC97254 [ Links ]
Snyman, H.A., Ingram, L.J. & Kirkman, K.P, 2013, 'Themeda triandra: a keystone grass species', African Journal of Range & Forage Science 30(3), 99-125. https://doi.org/10.2989/10220119.2013.831375 [ Links ]
South African National Biodiversity Institute, 2017, Red List of South African Plants version 2017.1, viewed 1 June 2019, from http://redlist.sanbi.org/ [ Links ]
Statistica, 2017, Statistica, version 13, TIBCO Software Inc., United States of America. [ Links ]
Veldman, J.W., Buisson, E., Durigan, G., Fernandes, G.W., Le Stradic, S., Mahy, G., Negreiros, D., Overbeck, G.E., Veldman, R.G., Zaloumis, N.P. & Putz, F.E., 2015, 'Toward an old growth concept for grasslands, savannas, and woodlands', Frontiers in Ecology and the Environment 13(3), 154-162. https://doi.org/10.1890/140270 [ Links ]
Zavaleta, E.S., Pasari, J.R., Hulvey, K.B. & Tilman, G.D., 2010, 'Sustaining multiple ecosystem functions in grassland communities requires higher biodiversity', Proceedings of the National Academy of Sciences 107(4), 1443-1446. https://doi.org/10.1073/pnas.0906829107 [ Links ]
 Correspondence:
Correspondence:
Prof. S.J. Siebert,
stefan.siebert@nwu.ac.za
Submitted: 20 September 2019
Accepted: 1 December 2020
Published: 24 February 2021


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}