










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.47 n.2 Pretoria 2017
http://dx.doi.org/10.4102/abc.v47i2.2143
ORIGINAL RESEARCH
Terrestrial invasions on sub-Antarctic Marion and Prince Edward Islands
Michelle GreveI; Rabia MathakuthaI; Christien SteynI; Steven L. ChownII
IDepartment of Plant and Soil Sciences, University of Pretoria, South Africa
IISchool of Biological Sciences, Monash University, Australia
ABSTRACT
BACKGROUND: The sub-Antarctic Prince Edward Islands (PEIs), South Africa's southernmost territories have high conservation value. Despite their isolation, several alien species have established and become invasive on the PEIs.
OBJECTIVES: Here we review the invasion ecology of the PEIs.
METHODS: We summarise what is known about the introduction of alien species, what influences their ability to establish and spread, and review their impacts.
RESULTS: Approximately 48 alien species are currently established on the PEIs, of which 26 are known to be invasive. Introduction pathways for the PEIs are fairly well understood - species have mainly been introduced with ship cargo and building material. Less is known about establishment, spread and impact of aliens. It has been estimated that less than 5% of the PEIs is covered by invasive plants, but invasive plants have attained circuminsular distributions on both PEIs. Studies on impact have primarily focussed on the effects of vertebrate invaders, of which the house mouse, which is restricted to Marion Island, probably has the greatest impact on the biodiversity of the islands. Because of the risk of alien introductions, strict biosecurity regulations govern activities at the PEIs. These are particularly aimed at stemming the introduction of alien species, and are likely to have reduced the rates of new introductions. In addition, some effort is currently being made to eradicate selected range-restricted species. However, only one species that had established and spread on the PEIs, the cat, has been successfully eradicated from the islands.
CONCLUSION: Given the ongoing threat of introductions, and the impacts of invaders, it is essential that future invasions to the PEIs are minimised, that the islands' management policies deal with all stages of the invasion process and that a better understanding of the risks and impacts of invasions is obtained.
Introduction
Islands are fragile ecosystems. Because of their isolation, islands are often taxonomically and functionally depauperate, that is, they house only a subset of the taxonomic or functional taxa compared to mainland ecosystem (Reaser et al. 2007). It is thought that this reduces biotic resistance to invasion and makes islands especially prone to invasions by alien species (Vitousek 1988; see also Gimeno, Vilà & Hulme 2006). Therefore, island species and ecosystems are thought to be particularly sensitive to the impacts of invasives (Donlan & Wilcox 2008; Reaser et al. 2007; see also Vilà et al. 2011). Despite better awareness of the risks of invasions and improved biosecurity regulations globally (García-de-Lomas & Vilà 2015; McGeoch et al. 2010), invasions continue unabated (Hulme 2009), with island invasions showing no sign of approaching a point of saturation (Sax & Gaines 2008).
Sub-Antarctic Islands represent some of the most isolated and least impacted habitats on the Earth; yet, they are also prone to invasions by nonindigenous species (Convey et al. 2006; Frenot et al. 2005; Shaw 2013). Indeed, alien invasions are thought to be the largest threat to the biodiversity of the sub-Antarctic Islands (Frenot et al. 2005), and a particularly high proportion of aliens that arrive on sub-Antarctic Islands appears to become invasive, of which many have negative impacts on native biodiversity (McGeoch et al. 2015).
The Prince Edward Islands (PEIs) constitute South Africa's southernmost, and only sub-Antarctic, territories (Figure 1). They consist of the larger Marion Island (46° 54' S, 37° 45' E; area: app. 270 km2) and the smaller Prince Edward Island (46° 38' S, 37° 57' E area: app. 45 km2), which are separated by approximately 19 km of ocean (Lutjeharms & Ansorge 2008). The islands are of volcanic origin, with an estimated age of 450 000 years (Boelhouwers et al. 2008). Marion Island reaches an altitude of 1230 m and Prince Edward Island an altitude of 672 m.
The PEIs have been a conservation area and a centre for scientific research since their occupation by South Africa in 1947. Along with other islands of the sub-Antarctic, the PEIs are of high conservation importance, where few small land masses support all terrestrial diversity in the region, including many of the top oceanic predators which use the islands as breeding grounds (de Villiers & Cooper 2008). In 1995, the PEIs were declared a Special Nature Reserve according to the Environment Conservation Act (No. 73 of 1989), which has now been superseded by the National Environmental Management: Protected Areas Act (No. 57 of 2003 NEMPA). Given the rich history of scientific activities on the PEIs (summarised in Chown & Froneman 2008b), their setting in a climatically dynamic area of the Southern Ocean (Fraser et al. 2009) and the fairly low anthropogenic pressures on the islands, the PEIs offer a valuable opportunity for studying and understanding ecological processes and management and research challenges, and how these challenges can be managed and forecast.
A sizeable number of alien species have been introduced to the PEIs (Table 1) (Chown & Froneman 2008b). Currently, approximately 18 (3) terrestrial plants, one (0) terrestrial vertebrate, 28 (5) terrestrial invertebrates and one (0) terrestrial fungus are thought to be established on Marion (and Prince Edward) Islands (Table 1). Some of the introduced species have a restricted distribution and have little effect on the native biodiversity on the islands (Gremmen 1997; Huntley 1971), but several (6 plants, 1 vertebrate and 19 invertebrates) have become invasive (invasion status D2 or E, Table 1) on the islands. (These are conservative estimates as the invasion status of other alien species is unknown, Table 1). Many of the invasives severely impact the native biodiversity and the ecosystem functioning of the islands. Indeed, along with climate change, invasive species probably pose the greatest conservation risk to the PEIs.
While a number of reviews about invasions to the greater sub-Antarctic region have been written (e.g. Convey et al. 2006; Frenot et al. 2005; Hughes & Convey 2010; Shaw 2013), we aim to review the state of knowledge of invasions for the PEIs. We specifically focus on invasions of terrestrial habitats, although the islands are also at risk of marine invasions (e.g. Lee & Chown 2007). The main aims of this paper are (1) to review what is known about alien and invasive organisms on the islands at different stages of the invasion process (Figure 2) (Blackburn et al. 2011). The first stage, introduction, deals with the arrival of organisms at the islands. In the second stage, establishment, we discuss how the alien species survive the environmental conditions on the island, grow and reproduce. Finally, the third stage comprises the spread of alien species, when they extend their ranges from the initial point of introduction to spread and occupy a larger range. (2) To summarise the known impacts of alien organisms. (3) To provide lists of the terrestrial alien species on the islands and what is known about their introduction, invasion status and impact. This information is especially relevant to South Africa's National Status Report on Biological Invasions, which will be released in 2017. (4) In light of what is known about invasions to the islands, we comment on management policies and current and future challenges for the PEIs.
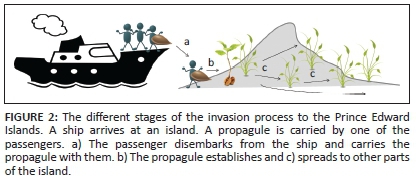
Human activities on the PEIs
Introductions of alien species to the PEIs are closely linked to the human activities on the islands. It is thought that the PEIs were first discovered in March 1663, but it was to be more than 100 years before the first person set foot on the islands. By 1802 the first sealers had made temporary camps on the islands (Gremmen 1975). Sealing activities continued on the island for approximately 50 years, during which time sealers were left to hunt seals on the islands for months at a time (Cooper 2008). After the seal populations had been decimated, human visits were relatively infrequent until 1947 when the islands were annexed by the South African government (Cooper 2008). Since annexation, Marion Island has been continuously inhabited by a small, though variable-sized (most recently in the old base approximately 15, since the building of the new base around 20), team of scientists and support staff, with no permanent residents living on the island (the maximum amount of time usually spent on the island is 13 months, although in exceptional circumstances individuals have stayed for 2 years). Prince Edward Island is unoccupied and only rarely visited. Due to the absence of a resident human population on the PEIs, the impact of humans has been limited compared to similar islands in the sub-Antarctic region (Chown, Gremmen & Gaston 1998). Nevertheless, human activities on Marion Island have been increasing steadily since the establishment of a permanent meteorological station on Marion Island in 1948. These anthropogenic activities have negatively impacted ecosystems and species on the island, with the introduction of alien or nonindigenous species being one particularly detrimental consequence (Chown & Froneman 2008a).
Human activities have been responsible for the introduction to the PEIs of a range of species from a variety of taxonomic groups (Chown & Froneman 2008b). The first alien introductions to the islands were associated with sealing vessels in the early 1800s. Subsequent military and scientific expeditions resulted in further introductions (Cooper 2008). Of all animal introductions in the PEIs, none have had a more substantial impact on the island's flora and fauna than the house mouse Mus musculus L. and the feral cat Felis catus L. The house mouse was accidentally introduced to Marion Island by sealers (Cooper 2008). Because of its tolerance of the sub-Antarctic climate and the absence of natural enemies on Marion Island, the house mouse was able to thrive on an omnivorous diet of plants and animals (Avenant & Smith 2003; Smith, Avenant & Chown 2002). In 1949, domestic cats were introduced to Marion Island in an attempt to deal with the mouse problem at the research station. These cats soon became feral, consuming large numbers of burrowing seabirds (Bester et al. 2002). They were also responsible for the extirpation of the common diving petrel (Pelecanoides urinatrix Gmelin) in 1965 (Bester et al. 2002). The cats were successfully removed in 1991, following a protracted eradication programme (Bester et al. 2002).
As human activities on the PEIs have increased over the years, so have the number of alien species and their impacts, and we are still coming to understand the invasion process and the impacts of invasives on the PEIs.
The stages of invasion
Introduction
The first stage of the invasion pathway is the introduction stage, when an organism is transported to an area to which it is non-native (Blackburn et al. 2011). Although many species that are introduced to areas outside their native range do not become invasive, some will do. Usually the greater the propagule pressure of non-native species to an area, the greater the chance that one of these species will become invasive (Simberloff et al. 2013). Therefore, the most effective way of curbing new alien introductions to an area is by gaining a thorough understanding of introduction pathways and vectors and using this information to inform the implementation of appropriate management actions and regulations to prevent future invasions (Faulkner et al. 2016; Hulme 2009; Keller & Kumschick 2017; Simberloff et al. 2013).
Islands are discreet landmasses; therefore, introduction pathways and the movement of vectors of alien species (i.e. people and goods) can often be effectively quantified (e.g. Christy, Savidge & Rodda 2007; Frenot, Gloaguen & Massé 2001). Likewise, the transport of goods and people to the PEIs is limited and well documented (DEA: Natural Resources Management Programme et al. 2012). In addition, there has been long-term and ongoing scientific research on the islands, with a recent focus on introduction pathways for the islands (e.g. Lee & Chown 2009a, 2009b, 2011). As a consequence, a fair amount is known about the timing, determinants and pathways of invasions to the PEIs.
An important determinant of alien introductions to an area is the nature of human activities the area is exposed to. Across the sub-Antarctic Islands, invasions are strongly influenced by the intensity of human activities: islands with more human visitors tend to have more introduced species (Chown et al. 2009; Frenot et al. 2005). The PEIs have no permanent human settlements; activities on the islands are restricted to scientific research and supporting activities, the protection or management of the islands and to reporting and educational activities, all of which are strictly regulated (Department of Environmental Affairs Directorate: Antarctica and Islands 2010). In addition, Marion Island is usually visited only once annually by one vessel. The current vessel transports approximately 80 scientists and support staff to the island, while Prince Edward Island is visited infrequently - at the most once every 4 years, and then only by a team of up to 10 scientists for a maximum duration of 8 days (Cooper et al. 2009; Department of Environmental Affairs Directorate: Antarctica and Islands 2010). Because visits to the PEIs are relatively infrequent, fewer alien species have been introduced to the islands compared to similar sub-Antarctic Islands with permanent settlements or more visitors (after controlling for climate, Chown et al. 1998). In addition, fewer alien species occur on the less visited Prince Edward Island than on Marion Island (de Villiers & Cooper 2008).
Aliens on the PEIs arrived at the islands via ships, their cargo and their occupants. A number of introductions occurred before the annexation of the PEIs, probably at the hands of sealers (Table 1, le Roux et al. 2013) although most introductions happened shortly after annexation in 1947/1948 (de Villiers & Cooper 2008). The building of the first research station in the 1960s coincided with a wave of new species being introduced to Marion Island (Huntley 1971; le Roux et al. 2013). Although existing biosecurity measures (see below) have been effective at reducing rates of introduction (see le Roux et al. 2013 for dates of discovery of alien plants on the PEIs), they have not prevented new species from arriving to the region.
Introductions to the islands have been intentional (e.g. cats) and unintentional (e.g. the birdeye pearlwort Sagina procumbens L., Figure 3a). Intentional introductions took place in the earlier years of the islands' occupation and included several vertebrate species. For example, the domestic pig (Sus scrofa L.) was deliberately introduced to Marion Island in 1804 where it bred successfully until it was eradicated some years later (Cooper 2008), while sheep, chickens and trout were introduced after annexation to provide fresh food to the overwintering teams (de Villiers & Cooper 2008; Gremmen 1975). None of these species have persisted. The only intentionally introduced species to the island that significantly impacted the native biodiversity was the cat (Cooper 2008; van Aarde 1980).
Some plant species were also intentionally introduced to the PEIs. After annexure, an attempt at planting several tree species, mostly of Northern hemisphere origin, was made; however, the conditions on the island (possibly the soils) proved unsuitable for their establishment (Gremmen 1975; La Grange 1954).
Most aliens were introduced accidentally. Introduction pathways are likely to have varied from rodents probably escaping off ships (Cooper 2008) to propagules being brought in clothing or cargo (Lee & Chown 2009a, 2009b, 2011), with building material (Bergstrom & Smith 1990; Slabber & Chown 2002), or in fresh produce (Smith 1992). During recent searches of cargo containers, passenger luggage and passenger clothing on voyages from Cape Town to Marion Island, live insects and plant seeds were retrieved (Lee & Chown 2009a, 2009b, 2011). The container packing location seemed to be a major source of propagules found in containers, and field clothing and field luggage contained a significant number of alien seeds (Lee & Chown 2009a).
Alien species are typically weedy Holarctic species which are also established in South Africa, from where the vessels visiting the PEIs most often depart (Lee & Chown 2011). However, because several scientists working on Marion Island also travel more widely in the greater Antarctic region, intra-regional transport of propagules to Marion Island is also likely (Lee & Chown 2011). Other studies have shown that people travelling to the region often visit and work in other cold climatic regions (Chown et al. 2012; Lee & Chown 2011).
Despite increasingly stringent policies to reduce introductions of alien species to the PEIs (DEA: Natural Resources Management Programme et al. 2012), new introductions are continuing (Table 1). Unsuccessful establishments of alien species are difficult to detect, but several new establishments have been recorded since stringent biosecurity regulations were put in place in the mid-1990s (e.g. Lee & Chown 2016; Slabber & Chown 2002), suggesting that new species continued being introduced. As a consequence, the main pathway of introductions, the annual relief voyages, is now subject to even greater biosecurity protocol than previously (Department of Environmental Affairs Directorate: Antarctica and Islands 2010). No new arrivals have been recorded since 2010, but systematic surveys for aliens on Marion Island have fallen away in the last few years - therefore, it is not possible to ascertain whether the absence of new records is because of increased biosecurity efforts or because of lack of surveys.
Introduction of aliens to Marion Island is mainly because of direct introductions by humans (whether intentional or unintentional). However, it is likely that some introductions of alien species to Prince Edward Island have been the result of secondary transport of alien propagules by natural vectors, such as wind or birds, from Marion to Prince Edward Island (Gremmen & Smith 1999; Ryan, Smith & Gremmen 2003). For example, the first record of S. procumbens L. on Prince Edward Island was in an area of high seal and bird activity, some distance from where scientists usually land on their expeditions to the island, making it unlikely that humans were responsible for the introduction (Gremmen & Smith 1999).
Which species reach the sub-Antarctic Islands such as the PEIs may be determined by dispersal ability. Vagility influences the nestedness of the different alien taxa of the sub-Antarctic Islands (Greve et al. 2005); in other words, assemblages of groups that are most mobile are more nested than those of less mobile groups. This suggests that organisms with better vagility are more likely to reach more sub-Antarctic Islands, whereas the arrival of species with lower vagility on the islands may be more haphazard (Greve et al. 2005; Shaw et al. 2010). Nevertheless, some overlap in alien assemblage composition across taxa is present across different sub-Antarctic Islands despite the large distances between them, with many of the alien species composing European weedy species (Frenot et al. 2005; Shaw et al. 2010).
Establishment
After introduction, the next stage of the invasion sequence is establishment; an organism must be able to survive and be viable in its new environment (Blackburn et al. 2011). This means that the environmental conditions must be favourable for the organism and that the organism is not driven to extinction by predators or pathogens in its new habitat.
The establishment of alien species on the PEIs is less well understood than the introduction vectors and pathways.
The alien vascular plant species that have established on the PEIs tend to originate from plant families with weedy tendencies (Shaw 2013), indicating that species from specific taxa are predisposed to being successful establishment.
The environment also plays an important role in establishment. Species richness of aliens on sub-Antarctic Islands is strongly influenced by climate: more alien species occur on warmer islands (Chown et al. 1998; Chown, Hull & Gaston 2005). This suggests that temperature is a major limiting factor to the establishment of alien species. Indeed, most alien species on the PEIs originate from the similarly cold Northern hemisphere (Shaw et al. 2010), rather than from milder South Africa, where most ship voyages originate from. (It is, however, important to note that many of the weedy plants established on the PEIs occur in South Africa and are common at the wharf precinct in Cape Town - the departure point of the supply ship; Lee & Chown 2009a). In addition, on Marion Island, many of the alien taxa are found in the milder lowlands, rather than in the more climatically harsh high-altitude areas (Chown et al. 2013; le Roux et al. 2013). Moreover, Lee et al. (2009) showed that the abundance of the invasive slug, Deroceras panormitanum (Lessona and Pollonera), was most strongly influenced by microclimatic conditions, with slugs being most abundant in conditions that corresponded to their physiological tolerances (Lee et al. 2009).
In many instances, plant invasions are promoted by disturbances and nutrient enrichment (Pyšek & Richardson 2008; Richardson et al. 2000). On the PEIs, the main agents of disturbance and nutrient enrichment are seals and seabirds (Smith 1978), which disturb the environment by trampling and burrowing (some seabirds) and enrich the soils through manuring. Accordingly, the establishment of alien plants on Marion Island is greater in areas that are prone to disturbances and nutrient enrichment by seals (Figure 3b and c) (Haussmann et al. 2013; Huntley 1971), and it is common to find alien species such as Poa annua L. growing at the entrance to bird burrows in regions where the incidence of alien species is otherwise low or non-existent.
Species that establish on the PEIs must possess adaptations to survive the conditions on the islands. These include low temperatures and strong winds. One of the most aggressive invasive plants on Marion Island is the grass P. annua (le Roux et al. 2013). It has spread extensively across the greater sub-Antarctic region and is the only vascular plant that has established and spread on the Antarctic continent (Chwedorzewska et al. 2015; Shaw et al. 2010). Its success in the region has been attributed to its high levels of phenotypic plasticity, which allow it to survive a range of different conditions. For example, it possesses the ability to switch between a perennial and annual life cycle depending on environmental conditions (reviewed in Chwedorzewska et al. 2015). In another example, the cold tolerance and the rapid generation time of the moth Plutella xylostella L. are considered pre-adaptations of the species to successfully establish on Marion Island (Crafford & Chown 1990).
Alien species that become invasive often possess a different set of traits to non-invasive aliens and native species (Ordonez, Wright & Olff 2010; van Kleunen, Weber & Fischer 2010), suggesting that some traits promote invasiveness. Identifying what traits confer higher invasiveness assists in identifying which species may be pre-adapted to becoming established and invasive to an area. Little is known about the role that functional traits play in the establishment of alien species on the PEIs. This has only been explored qualitatively for individual species of the PEIs (Crafford 1986; van Aarde 1986). For example, van Aarde (1986) suggested that cats could successfully establish on Marion Island because of a selective advantage of the founder population. The founder group showed a high incidence of the dark-coloured phenotype, the proportion of which increased over time, suggesting that this phenotype is associated with a selective advantage (discussed in van Aarde 1986). Therefore, not only the presence of the dark-coloured phenotype and its associated advantages in the founder population but also the selection for this phenotype as the population went through successive breeding cycles on Marion Island is thought to have aided the establishment of cats (van Aarde 1986).
A comparison of the traits of invasive species and indigenous species could provide insights into whether the establishment success of invasives is because they possess novel traits (i.e. they occupy niches not occupied by indigenous species) or because of their pre-adaptations to conditions on the island (under which circumstances one would expect these species to have traits similar to those of indigenous species) (Tecco et al. 2010). Only one published study has undertaken a semblance of such a study (Pammenter, Drennan & Smith 1986), although it entailed a comparison of only one pair of alien and indigenous con-generics (Agrostis stolonifera L. and Agrostis magellanica Lam., respectively). Therefore, its findings that the invasive species assimilated CO2 more effectively at low photon flux densities but showed less investment in sclerophyllous leaf tissue than the indigenous species cannot be generalised. Preliminary findings of a more recent study indicate that invasive plant species on Marion Island occupy functional niches not occupied by native species (Mathakutha et al., unpublished).
Dispersal and spread
After establishment, a non-native species must spread and extend its range in the new environment before it is classified an invasive (Blackburn et al. 2011). After an alien organism has started spreading, it usually becomes difficult and expensive to effectively control (Simberloff et al. 2013). It is thus recommended that efforts to eradicate alien species should be initiated before they reach the spread stage (Hughes & Convey 2012). In addition, factors that promote spread need to be thoroughly understood in order to prioritise eradication efforts.
Not all alien species introduced to the PEIs have spread on the islands - a number of species remain range-restricted (Table 1). Not much is known about the mode of spread of alien organisms on the PEIs once they have established. The rate and geography of spread are best understood for plants. Of the alien plant species that have successfully established on Marion Island, six have spread extensively since first detection (Table 1) (le Roux et al. 2013). Four of these are grasses. Sagina procumbens is thought to have spread at a rate of more than 1.5 km and 2 km per year on Marion and Prince Edward Island, respectively.
The rate of spread of alien plants on the uninhabited and infrequently visited Prince Edward Island suggests that natural (i.e. non-human) vectors play a significant role in the spread of invasive species (Ryan et al. 2003). Most alien plants on the PEIs possess a wind dispersal mechanism, and there is evidence that wind is a major determinant for the direction in which species disperse on the island (Born et al. 2012; Chown & Avenant 1992; Hedding, Nel & Anderson 2015). However, birds may also act as dispersal vectors (Hughes & Convey 2012; Ryan et al. 2003). For example, P. annua is frequently found growing at the entrances to bird burrows, also at higher altitudes, where the plant most likely arrives on the feathers or legs of seabirds (Bergstrom & Smith 1990). In addition to seabirds, seals are thought to aid alien spread by providing suitable high nutrient habitats which are favoured by many of the alien plant species (Haussmann et al. 2013; Ryan et al. 2003). Finally, on Marion Island, the spread of alien plants is most certainly aided by humans. This is obvious from the distribution of some alien plants growing along frequently used paths and their occurrence around island huts frequented by researchers (Figure 3d) (Gremmen & Smith 1999; le Roux et al. 2013).
The role of dispersal ability in driving the spread of alien species is well illustrated by two invasive invertebrates on Marion Island. The winged wasp Aphidius matricariae Haliday spread around Marion Island at a rapid rate of between 3 km and 5 km year−1 (Lee & Chown 2016). In contrast, the invasive springtail Pogonognathellus flavescens (L.) spread at a slow rate and has not yet occupied most niches suitable to it because of dispersal limitation (Treasure & Chown 2013). This can be explained by the fact that, in the absence of obvious dispersal vectors, the species is limited by low rates of movement between suitable habitats (Treasure & Chown 2013).
Some alien species on the PEIs may currently still be in a lag phase; that is, their populations are fairly restricted and stationery in extent, but they may spread more extensively in the future (Ryan et al. 2003). Other species are still filling suitable niches that have been uninhabited (Treasure & Chown 2013). For example, in 2013, the range of Poa pratensis L. on Marion Island was known to extend between approximately Long Ridge in the north and Bullard Beach in the south. By 2016, the species had also been recorded at the huts at Kildalkey in the south and Mixed Pickle and Swartkops in the west of the island (Steyn, Mathakutha & Greve, pers. obs., 2015&2016), suggesting that the species is still spreading.
It is difficult to ascertain what percentage of the island is invaded. For animal species, this has been poorly quantified (with the exception of individual species, e.g. Treasure & Chown 2013). For plants, it was estimated in 2008 that the overall cover of alien plants on the island was below 5% (Gremmen & Smith 2008). Another study which recorded localities of alien vascular plants in half-minute (app. 926 × 635 m) grid cells showed that by 2011 alien plant species had been recorded in approximately 42% and 53% of grid cells on Marion and Prince Edward Islands, respectively (le Roux et al. 2013). The lower percentage recorded on Marion Island is attributed to the higher altitudes of the island, at which few to no vascular plants grow. However, on Marion Island, almost all high nutrient lowland areas are invaded by one or several alien plant species that have circuminsular distributions (Gremmen & Smith 2008; le Roux et al. 2013).
Impacts
Invasive species are often considered to be a threat to native diversity and ecosystems because of the impacts they have on said diversity. Therefore, it is essential to understand what underlies these impacts (Levine et al. 2003). For the sub-Antarctic region, the impacts of introduced vertebrates on the biodiversity of islands have been most extensively studied (Frenot et al. 2005; Shaw 2013). Nevertheless, impacts of invasives on most indigenous taxa, with the exception perhaps of seabirds, are not well understood (Table 1) (McGeoch et al. 2015). On the PEIs, the impacts of only a handful of aliens have been investigated (Table 1), with the greatest focus being given to the study of mouse invasions (also see review in Chown & Froneman 2008a). The impacts of mouse invasions are fairly well understood for not only seabird populations but also some invertebrates, plants and ecosystem functioning and structure. In contrast, the impact of most other invasive species has received little attention.
Impacts of vertebrate predators on native biota are particularly severe because these species fulfil functional roles otherwise absent from the islands (Convey 2011; Courchamp, Chapuis & Pascal 2003). Organisms indigenous to the islands evolved in the absences of vertebrate predators and are thus poorly suited to defend themselves against these predators (Convey 2011; Frenot et al. 2005). Although the diet of cats on Marion Island included some mice, it mainly comprised seabirds, especially burrowing petrels (van Aarde 1980; van Rensburg 1983). Indeed, before cats were eradicated from Marion Island, their impact on seabird populations, especially burrowing species, was particularly severe, leading to decreased breeding success and reductions in their populations (van Rensburg 1983), and the extinction of at least one species on Marion Island (Bester et al. 2002). Furthermore, although seabird populations have seen some recovery after the reduction and eventual extirpation of cat populations on Marion Island (Cooper & Fourie 1991; Cooper et al. 1995), cat impacts have been long-lasting. Populations of burrowing petrels, which were particularly impacted by cats on Marion Island, have not recovered two decades after the eradication of the cats (Cerfonteyn & Ryan 2016; Smith 2008).
The house mouse, which occurs only on Marion Island, is probably the best-researched invader of the PEIs. One of its most obvious effects on the Marion Island system is through the extensive burrows that the animals construct (Figure 3e) (Avenant & Smith 2003; Eriksson & Eldridge 2014; Phiri, McGeoch & Chown 2009). Mouse activity causes direct and indirect damage to plants. The most notable direct damage to plants is to the keystone cushion plant, Azorella selago Hook. f. (Figure 3f) (Phiri et al. 2009). Mice burrow into the cushions, with the damage to cushions ranging from partial damage through entrance burrows to complete excavation, and thus disintegration of cushions (Phiri et al. 2009). The microclimate experienced by the cushion plants is also impacted by burrowing activities (Eriksson & Eldridge 2014). In addition, mice consume significant quantities of seeds, especially of the sedge Uncinia compacta R. Br., which has suffered significant population declines (Chown & Smith 1993; Smith & Steenkamp 1990). The impact of burrows on plants may also be indirect. Estimates of the amount of sediment removed through mouse burrows range between 2.4 t ha−1 and 8.4 t ha−1, with the latter probably being the more accurate estimate (Eriksson & Eldridge 2014). The large amount of sediment movement affects soil microclimate, with excavated soils being drier and warmer, and surface stability, resulting in higher levels of erosion (Eriksson & Eldridge 2014). Such changes to the soils are likely to impact the establishment of seedlings and the stability of plant communities (Eriksson & Eldridge 2014; Gremmen 1981), with likely knock-on effects on invertebrate assemblages (Hugo et al. 2004).
Invertebrates make up the bulk of the diet of the house mouse on Marion Island (Smith et al. 2002). The impacts of mice on invertebrates have mainly been assessed by comparing invertebrate assemblages on Marion Island and Prince Edward Island (free from mice) and by using mouse exclosures. It has been estimated that mice on Marion Island consume up to 2% of the invertebrate standing stock daily, and that mice predation has driven changes in population densities and mating strategies of some invertebrate species and a decline in the body size of weevil prey species (Chown & Smith 1993; Crafford & Scholtz 1987; Treasure & Chown 2014). Such predation is considered to have far-reaching effects on nutrient cycling (Crafford 1990b; Smith & Steenkamp 1990). A short-term (4-year) mouse exclosure experiment failed to reveal a significant effect of predation on invertebrate densities, although this could be because of poor statistical power of the analyses (van Aarde, Ferreira & Wassenaar 2004). By contrast, analyses of mouse and invertebrate densities over 40 years revealed significant and substantial impacts of mouse predation on key invertebrate populations (up to two orders of magnitude decline) (McClelland et al. in review).
More recently, it has been shown that mice are predating on albatross chicks on Marion Island (Jones & Ryan 2010). This behaviour was first recorded in 2003, but appears to be increasing in extent (it has been observed in colonies across the island) and frequency, and is resulting in chick mortalities on Marion Island (Dilley et al. 2016). This may partially be a consequence of the eradication of cats from the island, resulting in increased mouse populations which then give rise to mouse predatory behaviour (Dilley et al. 2016; Jones & Ryan 2010; Wanless et al. 2007). Predation by mouse on neighbouring Gough Island, first documented not long before the first record of mouse predation on Marion Island, has resulted in extensive losses of albatross and petrel chicks, causing populations of some seabirds to become unsustainable (Wanless et al. 2007). Seabirds on sub-Antarctic Islands tend to show some degree of naivety, probably as a result of a lack of natural predators in their native breeding habitat, possibly making them especially prone to mouse predation.
On islands, invasive plants cause fewer extinctions than invasive vertebrates (Sax & Gaines 2008). Nonetheless, plant invaders often alter whole habitats, thereby reducing populations of native species and affecting ecosystem processes and function (Reaser et al. 2007). The effects of invasive plants on the native diversity of the PEIs are poorly studied. Invasives occur across most vegetation complexes on the island (Gremmen 1997), with their effects most pronounced at low altitudes and in areas with high nutrient input (Gremmen & Smith 2008). Most of the invasive plants reach dominance in one or more vegetation complexes, in effect smothering the native vegetation and negatively impacting the diversity and abundance of native plants (Gremmen 1997; Gremmen et al. 1998). In some vegetation types, this may be because the leaves of the invasive grasses decompose more slowly than those of the indigenous species; thus the dry leaves accumulate and, in effect, block out the light (Gremmen 1997). One of the only studies on the impacts of an invasive plant concluded that the invasive grass, A. stolonifera, does not pose any threats of extinction to native species of plants or insects, although it forms dense stands which negatively affects the abundance of many native species (Figure 3g) (Gremmen et al. 1998). However, A. stolonifera is leading to the loss of native plant communities and changes in abiotic conditions (most especially light availability) for the bryophytes that grow in these communities, thereby fundamentally changing community structure (Gremmen et al. 1998). Although the impacts of invasive plants on Prince Edward Island biodiversity are only partially understood, many of the Prince Edward Island plant invaders are known to be invasive elsewhere (McGeoch et al. 2015) where they negatively impact native biodiversity.
Most studies on the impacts of alien species have focussed on impacts on individual native taxa. However, the knock-on effect of some impacts is evident in taxa that are indirectly affected by alien species. The insectivorous common sheathbill, Chionis minor Hartlaub, has seen significant declines, apparently because of invertebrate predation by mice (Huyser, Ryan & Cooper 2000). Indirect impacts of alien species can also be because of changes in the function and structure of ecosystems (Raymond et al. 2011). Of all alien taxa currently present on Marion Island, mice have the most obvious impact on ecosystem structure and functioning. Ecosystem functioning is most heavily impacted through changes in nutrient cycling because of the predation of macroinvertebrates that are instrumental in the decomposition process of the island (Smith 2008; Smith & Steenkamp 1990). This results in reduced rates of mineralisation and concomitant changes in the formation of peats and soils, which, in turn, affects vegetation succession and thus vegetation structure (Smith 2008).
Although some alien decomposers are present on the island, they do not necessarily fulfil identical functions to the indigenous macroinvertebrates that are heavily preyed upon by mice. For example, the introduced slug D. panormitanum is a decomposer; however, it mineralises N at a considerably slower rate than a comparative indigenous decomposer, the moth Pringleophaga marioni Viette (Smith 2007). In addition, mice preferentially feed on the indigenous P. marioni over the introduced D. panormitanum. This means that the build-up and the quality of peat on Marion Island are fundamentally altered by the higher abundance of the slug than P. marioni (Smith 2007).
While the known impacts of alien species are predominantly negative, alien species can also perform important ecosystem functions. Work by Hänel and Chown (1998) indicates that on Marion Island the introduced chironomid midge Lymnophyes minimus Meigen, also a litter feeder, is partly fulfilling the role of the heavily predated P. marioni, a keystone litter decomposer on the island (Crafford 1990a).
Invasions and climate change
It has been suggested that the synergy between two or more extinction drivers can exacerbate the risks to biodiversity that one of these drivers would otherwise pose alone (Brook, Sodhi & Bradshaw 2008). On the PEIs, such a synergy, in this case between alien invasions and climate change, is likely. The PEIs have experienced significant decreases in rainfall and increases in temperatures since climate monitoring on Marion Island started in 1951 (le Roux & McGeoch 2008; Smith & Steenkamp 1990). Climate change, especially the amelioration of temperatures, is expected to increase the vulnerability of sub-Antarctic systems to alien invasions (Convey 2011; Frenot et al. 2005). Indeed, the range expansions of several alien plant species on Marion Island (Chown et al. 2013; le Roux et al. 2013) and increases in mouse populations and their impacts (Chown & Smith 1993) are thought to have been driven by a warmer climate. Furthermore, physiological experiments on several invertebrate taxa indicate that alien species often display higher fitness or physiological tolerances than indigenous species under the warmer and drier conditions that may be expected under climate change (e.g. Chown et al. 2007; Janion et al. 2010; Slabber et al. 2007). Thus, there is strong evidence that climate change will exacerbate invasions and their impacts on Marion Island: alien species benefit from a milder climate, while native species decline, either because there are fewer areas where they can survive or because they are outcompeted by alien species (Chown & Froneman 2008a).
Taxonomic biases and gaps in knowledge
Globally, a strong taxonomic bias in invasion research exists (Pyšek et al. 2008). This is also true for the PEIs (Table 1), where the impacts of vertebrates are best understood, followed by plants and insects (Table 1). For these taxa, the listed species (Table 1) present a fairly comprehensive account of aliens on the islands. However, other taxa have generally been poorly studied, and therefore little is known about species in these taxa. For example, while there is a high diversity of lichens on the islands (Øvstedal & Gremmen 2011), records of new species are still being added for the island (N.J. Gremmen, pers. comm., 2015). None of the lichens that have been identified on the PEIs are known to have been introduced by humans. Similarly, no alien algae have been described (Van de Vijver, Gremmen & Smith 2008; van Staden 2011), although there is evidence that these groups are being transported to the region (Huiskes et al. 2014).
Microbes are not visible, yet they are ubiquitous and easily transported organisms (Cowan et al. 2011). The microbiology of the PEIs is poorly known (Sanyika, Stafford & Cowan et al. 2012), but it is highly probable that a number of microbes have been introduced to the PEIs by humans (see discussion in Cowan et al. 2011; Hughes, Cowan & Wilmotte 2015). The only alien microbe known to have been introduced to the PEIs is the fungal pathogen Botryotina fuckeliana (de Bary) Whetzel on Marion Island (Kloppers & Smith 1998). It is thought to have been introduced in fresh produce and attacks the Kerguelen cabbage, Pringlea antiscorbutica R. Br. (Kloppers & Smith 1998). Other possible pathways by which microbes may have been introduced to the PEIs are diverse (Cowan et al. 2011) and could include footwear, scientific equipment, foods with yeasts, human excrement or the soils that were brought from South Africa to Marion Island for greenhouse experiments shortly after annexation (La Grange 1954). It is thus essential to understand what microbes are being introduced to the islands by which pathways, how readily they spread on the islands and what their impacts are.
Invasion policies and management
Globally, the territories of the Antarctic region, including the PEIs, have some of the strictest regulations governing introduced species (de Villiers et al. 2006; Hughes & Pertierra 2016). The South African Department of Environmental Affairs manages alien species on the PEIs, with most efforts directed at aliens on Marion Island which has more introduced species and a permanent scientific contingent.
The best way to minimise invasions and their impacts is to prevent them (Simberloff et al. 2013). Accordingly, the largest efforts in alien control in the Antarctic region are concentrated on reducing introductions. In contrast, monitoring and managing existing invasive plants receive less attention (McGeoch et al. 2015). This is also the case for the PEIs. The Prince Edward Islands Management Plan (Department of Environmental Affairs Directorate: Antarctica and Islands 2010) deals most stringently with the introduction stage of the invasion process. The management plan currently sets detailed regulations with the aim of preventing introductions of alien species to the islands. A stringent vessel inspection is required prior to departure to the islands, and the vessel has many trapping techniques deployed for animal (rodent and invertebrate) propagules. Likewise, cargo, personal equipment and clothing are all inspected prior to departure, en route to the island (Figure 3h), and on landing at the scientific station, where all cargo is delivered prior to field deployment to other parts of the island (Department of Environmental Affairs Directorate: Antarctica and Islands 2010). In addition, there is a ban on the import of organic materials, soils or rocks to the islands, and the release of ballast water or food waste from the supply vessel is prohibited in the proximity of the islands (Department of Environmental Affairs Directorate: Antarctica and Islands 2010). While these biosecurity regulations probably contribute to lower introduction rates to the PEIs, they are not always strictly enforced and do not entirely preclude the introduction of new species.
Once an alien species has arrived on the PEIs, it must be detected to be controlled. Detection of many species is dependent on scientists with knowledge of the taxa on the island spotting new species. Currently, no specific surveys for the purpose of detection take place; instead, detection is dependent on organisms being recorded during other scientific or monitoring activities. Identification can be aided by genetic methods: in cases where species identification is difficult, DNA barcoding or similar methods can be employed to aid identification of new organisms (Chown, Sinclair & van Vuuren 2008).
Some alien species, especially those first recorded at the base, have been eradicated from Marion Island upon first detection before they had the opportunity to establish (Hänel, Chown & Davies 1998). Other species have probably disappeared from the islands because they were not able to establish (e.g. Gremmen & Smith 1999), or through a combination of eradication efforts and climatic conditions unfavourable to the species (e.g. Cooper et al. 1992). Currently, there are ongoing efforts to eradicate some alien species on Marion Island (Table 2). These efforts are especially focussed on alien plants with restricted distributions, that is, species that have established but not spread extensively, with the preferred method of control being targeted herbicide use (DEA: Natural Resources Management Programme et al. 2012). Because they have restricted ranges and relatively low rates of dispersal, plants are easier to eradicate than most animal taxa. At present eradication efforts of restricted-range species are effective in preventing these species from spreading, and some species have been eradicated (Tables 1 and 2).
The eradication of species that have established and become widespread is difficult, and accordingly little management of these species takes place. Indeed, it has been argued that the removal of many plant species and most invertebrates from sub-Antarctic Islands such as the PEIs is impractical (Convey 2011). The eradication of cats from Marion Island constitutes the most ambitious attempt at eradicating a species on the island, as well as the only eradication of a widespread and established species. The biological control agent feline panleukopenia virus was introduced to Marion Island in 1977 to reduce cat populations. The remaining, but diminished, cat population was then exterminated through hunting (Bester et al. 2002). Interestingly, one other biological control agent was introduced to Marion Island accidentally, which has led to the inadvertent control of an invasive aphid. The wasp A. matricariae, a parasitoid of aphids, was accidentally introduced to Marion Island in cargo between 2001 and 2003 (Lee et al. 2007) and has rapidly spread across the island where it appears to be effectively controlling populations of the invasive aphid Rhopalosiphum padi (L.) (Lee & Chown 2016).
More recently, investigations into the feasibility of eradicating mice from Marion Island by chemical means have been conducted. Rodent eradications have been successful on other islands (Howald et al. 2007). Such an operation on Marion Island would be expensive and logistically complex given the difficult terrain and weather, but not impossible (e.g. Springer 2016).
Finally, while humans are the main agents of species introductions to the PEIs, it must be kept in mind that new species may arrive to the islands from their native range by natural means and that these species should thus not be the subject of control measures as they are colonisations (Lee, Terauds & Chown 2014). Examples of what are thought to be natural dispersal events to Marion Island include the globally invasive diamondback moth P. xylostella (Chown & Avenant 1992) and the shrub Ochetophila trinervis (Gillies ex Hook.) Poepp. ex Endl. (Gremmen & Smith 2008; Kalwij et al. unpublished data). To determine the mode of arrival of an alien species at the PEIs may be difficult. However, if a new species is first detected at a considerable distance from sites of high human activities on the islands, it may indicate that the species arrived by natural means (Chown & Avenant 1992; Gremmen & Smith 1999; Lee et al. 2014). In some cases, genetic tools may be used to narrow down the region of origin of the species, providing further clues as to whether the species arrived by natural or by human-mediated means (Chown & Avenant 1992; Lee et al. 2014). However, where uncertainty exists as to whether a species is a natural colonist, a precautionary approach has been recommended, where a species is eradicated before it spreads (Hughes & Convey 2012).
Implications for the national status report on biological invasions
The current pathways of human-assisted introductions of alien species to the PEIs are straightforward because all species arrive by ship. Thus, pathways can be narrowed down to unintentional introductions, mainly through cargo, building material and clothes. For those organisms that have established on the islands, the status of invasion is well known for plants and the one vertebrate on the island, but only for some insect species. Not all introduced species establish and spread, but those that do have spread extensively across both islands. The mouse has the largest impact on the biodiversity of the PEIs - impacts span trophic levels and affects island-scale ecosystem functioning. The impacts of invasive plants are thought to be high in some vegetation types, while they are negligible in others. In contrast, impacts of invertebrates are poorly understood.
Current control interventions of some range-restricted alien species on Marion Island are effective at preventing their populations from spreading, although interventions for widely distributed species are absent. No interventions are being implemented on Prince Edward Island, which has received a number of alien species by natural vectors from Marion Island.
In summary, Marion Island is fairly heavily impacted by alien species, with extensive effects recorded for native species and ecosystem functioning. Prince Edward Island is currently invaded by a comparatively small number of species. While no works on the impacts of invasive species have been conducted on Prince Edward Island, the impacts are likely to be significantly less than those on Marion Island on account of the number and the identity of the invasive species found on Prince Edward Island.
Future prospects and conclusion
Along with climate change, the introduction of alien species probably entails the greatest threat to the biodiversity of the PEIs. Despite existing biosecurity regulations, transport of nonindigenous propagules to the region continues (Houghton et al. 2016). Therefore, to ensure the continued conservation of the PEIs, it is of utmost importance that alien introductions and invasions are kept to a minimum. Prevention remains the easiest, and cheapest, way to keep alien species off the PEIs (Hughes & Pertierra 2016). It must thus be ensured that strict biosecurity regulations aimed at reducing propagule and live individual entrainment (Department of Environmental Affairs Directorate: Antarctica and Islands 2010) are implemented by the Department of Environmental Affairs. Furthermore, it is critical that these regulated biosecurity protocols are enforced and that participants and managers of voyages to the islands are aware of the requirements. In addition, while the focus of this article has been on terrestrial organisms, the threats of marine and, perhaps to a lesser extent, freshwater invasions must not be overlooked (Table 4-A1) (Dartnall & Smith 2012; Lee & Chown 2007).
While efforts at controlling range-limited species have managed to contain the spread of a number of alien species on Marion Island, regular field surveys are important for the detection and recording of new occurrences or recent range expansions of alien species. Such activities would be most effective around sites of high human activities to where alien species are most likely to be introduced and spread from (Hughes & Pertierra 2016; le Roux et al. 2013). This allows for early detection and the implementation of rapid response measures to contain spread (Hughes & Pertierra 2016; Huntley 1971). On Marion Island, regular surveys aimed at detecting new arrivals to the island were conducted between the early 1980s and 2011 (Lee & Chown 2016). These surveys no longer take place. If such surveys by field experts should be regularly undertaken and be a standard component of the formal management plan of the islands, it would provide better continuity.
A current challenge to undertaking such surveys is the recent loss of capacity of entomological and other invertebrate knowledge, and possibly the imminent loss of bryophyte knowledge, from the scientific community within the South African National Antarctic Programme (SANAP) community. A lack of capacity to identify species from these taxonomic groups can limit the ability to detect new arrivals to the islands. Thus, retention of taxonomic knowledge within the SANAP programme is essential for continued alien species management. Alternatively, efforts to build capacity should be supported, such as inviting international experts to train new cohorts of researchers.
In addition, monitoring of widespread alien species should be a priority to ensure that management policies incorporate control or mitigation measures where possible (Hughes & Pertierra 2016). Currently, monitoring activities of widespread aliens is conducted on an ad hoc basis every few years as part of the research programmes of researchers in SANAP (e.g. Gremmen 1975; le Roux et al. 2013; Ryan et al. 2003). A more formalised approach may be desirable.
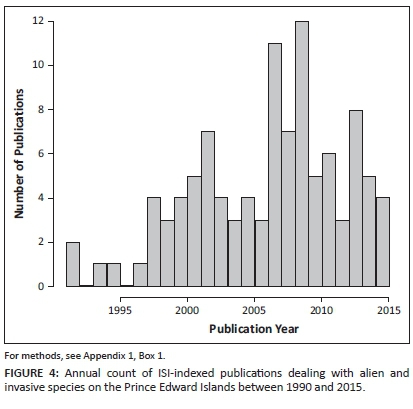
Finally, it is important that scientific studies on the extent and impacts of introduced species on the PEIs should continue. To effectively channel efforts for invasion management, it is essential to understand which invasive species pose the greatest threat to diversity, what their precise impacts are, and to identify in which systems such impacts are most pronounced (Gurevitch & Padilla 2004). Research on biological invasions on the PEIs has enjoyed steady attention in recent decades. A survey of a number of papers about alien and invasive species on the PEIs shows a steady increase from the early 1990s to the late 2000s, with a recent decline since 2009 (Figure 4). Given the ongoing threat of invasions to the PEIs, and the need to better understand and manage their impacts, it is imperative that the invasion biology of the PEIs remains an important focus of research on the islands. For example, a risk assessment of which species might become invasive would assist in preventing future invasions. Such risk assessments can be based on climatic suitability or on species traits known to be predictors of invasive capability. While the introduction stage of invasions is reasonably well understood for the PEIs, less attention has been given to the process of alien establishment and spread. In addition, the impacts of many of the alien species need to be better understood. Finally, basic science on the biology of native species and ecosystem processes on the islands needs to continue. Without such an understanding, the impacts of invasions cannot be fully appreciated.
Acknowledgements
Two anonymous reviewers provided helpful comments on earlier versions of this review. The authors thank Peter le Roux for discussions.
The South African NRF-funded National Antarctic Programme is thanked for providing financial support to M.G. (Grant No. 93065).
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Authors' contributions
M.G. led the writing, with contributions from R.M., C.S. and S.L.C. M.G., R.M. and C.S. collated information on current invasions with input from S.L.C.
References
Avenant, N.L. & Smith, V.R., 2003, 'The microenvironment of house mice on Marion Island (sub-Antarctic)', Polar Biology 26(2), 129-141. [ Links ]
Bergstrom, D.M. & Smith, V.R., 1990, 'Alien vascular flora of Marion and Prince Edward Islands: New species, present distribution and status', Antarctic Science 2(4), 301-308. https://doi.org/10.1017/S0954102090000426 [ Links ]
Bester, M.N., Bloomer, J.P., van Aarde, R.J., Erasmus, B.H., van Rensburg, P.J.J., Skinner, J.D. et al., 2002, 'A review of the successful eradication of feral cats from sub-Antarctic Marion Island, Southern Indian Ocean', South African Journal of Wildlife Research 32(1), 65-73. [ Links ]
Blackburn, T.M., Essl, F., Evans, T., Hulme, P.E., Jeschke, J.M., Kühn, I. et al., 2014, 'A unified classification of alien species based on the magnitude of their environmental impacts', PLoS Biology 12(5), e1001850. https://doi.org/10.1371/journal.pbio.1001850 [ Links ]
Blackburn, T.M., Pyšek, P., Bacher, S., Carlton, J.T., Duncan, R.P., Jarošík, V. et al., 2011, 'A proposed unified framework for biological invasions', Trends in Ecology & Evolution 26(7), 333-339. https://doi.org/10.1016/j.tree.2011.03.023 [ Links ]
Boelhouwers, J.C., Meiklejohn, K.I., Holness, S.D. & Hedding, D.W., 2008, 'Geology, geomorphology and climate change', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands: Land-sea interactions in a changing ecosystem, pp. 65-96, SUN PReSS, Stellenbosch. [ Links ]
Born, C., le Roux, P.C., Spohr, C., McGeoch, M.A. & Jansen van Vuuren, B., 2012, 'Plant dispersal in the sub-Antarctic inferred from anisotropic genetic structure', Molecular Ecology 21(1), 184-194. https://doi.org/10.1111/j.1365-294X.2011.05372.x [ Links ]
Brook, B.W., Sodhi, N.S. & Bradshaw, C.J.A., 2008, 'Synergies among extinction drivers under global change', Trends in Ecology & Evolution 23(8), 453-460. https://doi.org/10.1016/j.tree.2008.03.011 [ Links ]
Cerfonteyn, M. & Ryan, P.G., 2016, 'Have burrowing petrels recovered on Marion Island two decades after cats were eradicated? Evidence from sub-Antarctic skua prey remains', Antarctic Science 28(1), 51-57. https://doi.org/10.1017/S0954102015000474 [ Links ]
Chown, S.L. & Avenant, N., 1992, 'Status of Plutella xylostella at Marion Island six years after its colonisation', South African Journal of Antarctic Research 22(1&2), 37-40. [ Links ]
Chown, S.L. & Froneman, P.W., 2008a, 'Conclusion: Change in terrestrial and marine systems', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands: Land-sea interactions in a changing ecosystem, pp. 351-372, African SunMedia, Stellenbosch. [ Links ]
Chown, S.L. & Froneman, P.W. (eds.), 2008b, The Prince Edward Islands: Land-Sea Interactions in a Changing Ecosystem, African SunMedia, Stellenbosch. [ Links ]
Chown, S.L., Gremmen, N.J.M. & Gaston, K.J., 1998, 'Ecological biogeography of southern ocean islands: Species-area relationships, human impacts, and conservation', American Naturalist 152(4), 562-575. https://doi.org/10.1086/286190 [ Links ]
Chown, S.L., Huiskes, A.H.L., Gremmen, N.J.M., Lee, J.E., Terauds, A., Crosbie, K. et al., 2012, 'Continent-wide risk assessment for the establishment of nonindigenous species in Antarctica', Proceedings of the National Academy of Sciences of the United States of America 109(13), 4938-4943. https://doi.org/10.1073/pnas.1119787109 [ Links ]
Chown, S.L., Hull, B. & Gaston, K.J., 2005, 'Human impacts, energy availability and invasion across Southern Ocean Islands', Global Ecology and Biogeography 14(6), 521-528. https://doi.org/10.1111/j.1466-822x.2005.00173.x [ Links ]
Chown, S.L., le Roux, P.C., Ramaswiela, T., Kalwij, J.M., Shaw, J.D. & McGeoch, M.A., 2013, 'Climate change and elevational diversity capacity: Do weedy species take up the slack?', Biology Letters 9(1), 20120806. https://doi.org/10.1098/rsbl.2012.0806 [ Links ]
Chown, S.L., Sinclair, B.J. & Vuuren, B.J., 2008, 'DNA barcoding and the documentation of alien species establishment on sub-Antarctic Marion Island', Polar Biology 31(5), 651-655. https://doi.org/10.1007/s00300-007-0402-z [ Links ]
Chown, S.L., Slabber, S., McGeoch, M.A., Janion, C. & Leinaas, H.P., 2007, 'Phenotypic plasticity mediates climate change responses among invasive and indigenous arthropods', Proceedings of the Royal Society B: Biological Sciences 274(1625), 2531-2537. https://doi.org/10.1098/rspb.2007.0772 [ Links ]
Chown, S.L. & Smith, V.R., 1993, 'Climate change and the short-term impact of feral house mice at the sub-Antarctic Prince Edward Islands', Oecologia 96(4), 508-516. https://doi.org/10.1007/BF00320508 [ Links ]
Chown, S.L., Spear, D., Lee, J.E. & Shaw, J.D., 2009, 'Animal introductions to southern systems: Lessons for ecology and for policy', African Zoology 44(2), 248-262. https://doi.org/10.3377/004.044.0213 [ Links ]
Christy, M.T., Savidge, J.A. & Rodda, G.H., 2007, 'Multiple pathways for invasion of anurans on a Pacific island', Diversity and Distributions 13(5), 598-607. https://doi.org/10.1111/j.1472-4642.2007.00378.x [ Links ]
Chwedorzewska, K.J., Gielwanowska, I., Olech, M., Molina-Montenegro, M.A., Wódkiewicz, M. & Galera, H., 2015, 'Poa annua L. in the maritime Antarctic: An overview', Polar Record 51(6), 637-643. https://doi.org/10.1017/S0032247414000916 [ Links ]
Convey, P., 2011, 'Antarctic terrestrial biodiversity in a changing world', Polar Biology 34(11), 1629-1641. https://doi.org/10.1007/s00300-011-1068-0 [ Links ]
Convey, P., Frenot, Y., Gremmen, N.J.M. & Bergstrom, D.M., 2006, 'Biological invasions', in D.M. Bergstrom, P. Convey & A.H.L. Huiskes (eds.), Trends in Antarctic terrestrial and limnetic ecosystems: Antarctica as a global indicator, pp. 193-220, Springer, Dordrecht. [ Links ]
Cooper, J., 2008, 'Human history', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands: Land-sea interactions in a changing ecosystem, pp. 331-350, SUN PReSS, Stellenbosch. [ Links ]
Cooper, J., Bester, M.N., Chown, S.L., Crawford, R.J.M., Daly, R., Heyns, E. et al., 2009, 'Biological survey of the Prince Edward Islands, December 2008', South African Journal of Science 105(7&8), 317-320. [ Links ]
Cooper, J., Crafford, J.E. & Hecht, T., 1992, 'Introduction and extinction of brown trout (Salmo trutta L.) in an impoverished subantarctic stream', Antarctic Science 4(1), 9-14. https://doi.org/10.1017/S095410209200004X [ Links ]
Cooper, J. & Fourie, A., 1991, 'Improved breeding success of great-winged petrels Pterodroma macroptera following control of feral cats Felis catus at subantarctic Marion Island', Bird Conservation International 1(2), 171-175. https://doi.org/10.1017/S0959270900002033 [ Links ]
Cooper, J., Marais, A.V.N., Bloomer, J.P. & Bester, M.N., 1995, 'A success story: Breeding of burrowing petrels (Procellariidae) before and after the eradication of feral cats Felis catus at subantarctic Marion Island', Marine Ornithology 23(1), 33-37. [ Links ]
Courchamp, F., Chapuis, J.L. & Pascal, M., 2003, 'Mammal invaders on islands: Impact, control and control impact', Biological Reviews 78(3), 347-383. https://doi.org/10.1017/S1464793102006061 [ Links ]
Cowan, D.A., Chown, S.L., Convey, P., Tuffin, M., Hughes, K., Pointing, S. et al., 2011, 'Non-indigenous microorganisms in the Antarctic: Assessing the risks', Trends in Microbiology 19(11), 540-548. https://doi.org/10.1016/j.tim.2011.07.008 [ Links ]
Crafford, J.E., 1986, 'A case study of an alien invertebrate (Limnophyes pusillus, Diptera, Chironomidae) introduced on Marion Island: Selective advantages', South African Journal of Antarctic Research 16(3), 115-117. https://doi.org/10.1016/0006-3207(87)90119-4 [ Links ]
Crafford, J.E., 1990a, 'Patterns of energy flow in populations of the dominant insect consumers on Marion Island', DPhil thesis, Department of Entomology, University of Pretoria. [ Links ]
Crafford, J.E., 1990b, 'The role of feral house mice in ecosystem functioning on Marion Island', in K.R. Kerry & G. Hempel (eds.), Antarctic ecosystems. Ecological change and conservation, pp. 359-364, Springer-Verlag, Berlin. [ Links ]
Crafford, J.E. & Chown, S.L., 1990, 'The introduction and establishment of the diamondback moth (Plutella xylostella L., Plutellidae) on Marion Island', in K.R. Kerry & G. Hempel (eds.), Antarctic ecosystems. Ecological change and conservation, pp. 354-358, Springer-Verlag, Berlin. [ Links ]
Crafford, J.E. & Scholtz, C.H., 1987, 'Quantitative differences between the insect faunas of Sub-Antarctic Marion and Prince Edward Islands: A result of human intervention?', Biological Conservation 40(4), 255-262. [ Links ]
Dartnall, H.J.G. & Smith, V.R., 2012, 'Freshwater invertebrates of sub-Antarctic Marion Island', African Zoology 47(2), 203-215. https://doi.org/10.3377/004.047.0207 [ Links ]
de Villiers, M.S. & Cooper, J., 2008, 'Conservation and management', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands: Land-sea interactions in a changing ecosystem, pp. 301-330, SUN PReSS, Stellenbosch. [ Links ]
de Villiers, M.S., Cooper, J., Carmichael, N., Glass, J.P., Liddle, G.M., Mclvor., E. et al., 2006, 'Conservation management at Southern Ocean Islands: Towards the development of best-practice guidelines', Polarforschung 75(2/3), 113-131. [ Links ]
DEA: Natural Resources Management Programme, DEA: Southern Oceans and Antarctic Support, DEA: Environmental Impact Evaluation & DST-NRF Centre of Excellence for Invasion Biology: Stellenbosch University, 2012, Eradication, monitoring and control of alien and invasive alien species on Marion Island, Unpublished. [ Links ]
Department of Environmental Affairs Directorate: Antarctica and Islands, 2010, Prince Edward Islands Management Plan Version 0.2, Department of Environmental Affairs, Directorate: Antarctica and the Islands. [ Links ]
Dilley, B.J., Schoombie, S., Schoombie, J. & Ryan, P.G., 2016, '"Scalping" of albatross fledglings by introduced mice spreads rapidly at Marion Island', Antarctic Science 28(2), 73-80. https://doi.org/10.1017/S0954102015000486 [ Links ]
Donlan, C.J. & Wilcox, C., 2008, 'Diversity, invasive species and extinctions in insular ecosystems', Journal of Applied Ecology 45(4), 1114-1123. https://doi.org/10.1111/j.1365-2664.2008.01482.x [ Links ]
Eriksson, B. & Eldridge, D.J., 2014, 'Surface destabilisation by the invasive burrowing engineer Mus musculus on a sub-Antarctic island', Geomorphology 223, 61-66. https://doi.org/10.1016/j.geomorph.2014.06.026 [ Links ]
Faulkner, K.T., Robertson, M.P., Rouget, M. & Wilson, J.R.U., 2016, 'Understanding and managing the introduction pathways of alien taxa: South Africa as a case study', Biological Invasions 18(1), 73-87. https://doi.org/10.1007/s10530-015-0990-4 [ Links ]
Fraser, C.I., Nikula, R., Spencer, H.G. & Waters, J.M., 2009, 'Kelp genes reveal effects of subantarctic sea ice during the Last Glacial Maximum', Proceedings of the National Academy of Sciences 106(9), 3249-3253. https://doi.org/10.1073/pnas.0810635106 [ Links ]
Frenot, Y., Chown, S.L., Whinam, J., Selkirk, P.M., Convey, P., Skotnicki, M. et al., 2005, 'Biological invasions in the Antarctic: Extent, impacts and implications', Biological Reviews 80(1), 45-72. https://doi.org/10.1017/S1464793104006542 [ Links ]
Frenot, Y., Gloaguen, J.-C. & Massé, L., 2001, 'Human activities, ecosystem disturbances and plant invasions in the French islands of the southern Indian Ocean (Crozet, Kerguelen and Amsterdam Islands)', Biological Conservation 101(1), 33-50. https://doi.org/10.1016/S0006-3207(01)00052-0 [ Links ]
García-de-Lomas, J. & Vilà, M., 2015, 'Lists of harmful alien organisms: Are the national regulations adapted to the global world?', Biological Invasions 17(11), 3081-3091. https://doi.org/10.1007/s10530-015-0939-7 [ Links ]
Gimeno, I., Vilà, M. & Hulme, P.E., 2006, 'Are islands more susceptible to plant invasion than continents? A test using Oxalis pes-caprae L. in the western Mediterranean', Journal of Biogeography 33(9), 1559-1565. https://doi.org/10.1111/j.1365-2699.2006.01525.x [ Links ]
Gremmen, N.J.M., 1975, 'The distribution of alien vascular plant on Marion and Prince Edward Islands', South African Journal of Antarctic Research 5, 25-30. [ Links ]
Gremmen, N.J.M., 1981, The vegetation of the Subantarctic islands Marion and Prince Edward, Dr. W. Junk, The Hague. [ Links ]
Gremmen, N.J.M., 1997, 'Changes in the vegetation of sub-Antarctic Marion Island resulting from introduced vascular plants', in B. Battaglia, J. Valencia & D.W.H. Walton (eds.), Antarctic communities: Species, structure and survival, pp. 417-423, Cambridge University Press, Cambridge. [ Links ]
Gremmen, N.J.M., Chown, S.L. & Marshall, D.J., 1998, 'Impact of the introduced grass Agrostis stolonifera on vegetation and soil fauna communities at Marion Island, sub-Antarctic', Biological Conservation 85(3), 223-231. https://doi.org/10.1016/S0006-3207(97)00178-X [ Links ]
Gremmen, N.J.M. & Smith, V.R., 1999, 'New records of alien vascular plants from Marion and Prince Edward Islands, Sub-Antarctic', Polar Biology 21(6), 401-409. https://doi.org/10.1007/s003000050380 [ Links ]
Gremmen, N.J.M. & Smith, V.R., 2008, 'Terrestrial vegetation and dynamics', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands: Land-sea interactions in a changing ecosystem, pp. 215-244, African SunMedia, Stellenbosch. [ Links ]
Greve, M., Gremmen, N.J.M., Gaston, K.J. & Chown, S.L., 2005, 'Nestedness of Southern Ocean island biotas: Ecological perspectives on a biogeographical conundrum', Journal of Biogeography 32(1), 155-168. https://doi.org/10.1111/j.1365-2699.2004.01169.x [ Links ]
Gurevitch, J. & Padilla, D.K., 2004, 'Are invasive species a major cause of extinctions?', Trends in Ecology & Evolution 19(9), 470-474. https://doi.org/10.1016/j.tree.2004.07.005 [ Links ]
Hänel, C. & Chown, S.L., 1998, ''The impact of a small, alien invertebrate on a sub-Antarctic terrestrial ecosystem: Limnophyes minimus (Diptera, Chironomidae) at Marion Island', Polar Biology 20(2), 99-106. https://doi.org/10.1007/s003000050282 [ Links ]
Hänel, C., Chown, S.L. & Davies, L., 1998, 'Records of alien insect species from sub-Antarctic Marion and South Georgia Islands', African Entomology 6(2), 366-369. [ Links ]
Haussmann, N.S., Rudolph, E.M., Kalwij, J.M. & McIntyre, T., 2013, 'Fur seal populations facilitate establishment of exotic vascular plants', Biological Conservation 162, 33-40. https://doi.org/10.1016/j.biocon.2013.03.024 [ Links ]
Hedding, D.W., Nel, W. & Anderson, R.L., 2015, 'Aeolian processes and landforms in the sub-Antarctic: Preliminary observations from Marion Island', Polar Research 34, 26365. https://doi.org/10.3402/polar.v34.26365 [ Links ]
Houghton, M., McQuillan, P., Bergstrom, D., Frost, L., van den Hoff, J. & Shaw, J., 2016, 'Pathways of alien invertebrate transfer to the Antarctic region', Polar Biology 39(1), 23-33. https://doi.org/10.1007/s00300-014-1599-2 [ Links ]
Howald, G., Donlan, C.J., Galván, J.P., Russell, J.C., Parkes, J., Samaniego, A. et al., 2007, 'Invasive rodent eradication on islands', Conservation Biology 21(5), 1258-1268. https://doi.org/10.1111/j.1523-1739.2007.00755.x [ Links ]
Hughes, K.A. & Convey, P., 2010, 'The protection of Antarctic terrestrial ecosystems from inter- and intra-continental transfer of non-indigenous species by human activities: A review of current systems and practices', Global Environmental Change 20(1), 96-112. https://doi.org/10.1016/j.gloenvcha.2009.09.005 [ Links ]
Hughes, K.A. & Convey, P., 2012, 'Determining the native/non-native status of newly discovered terrestrial and freshwater species in Antarctica - Current knowledge, methodology and management action', Journal of Environmental Management 93(1), 52-66. https://doi.org/10.1016/j.jenvman.2011.08.017 [ Links ]
Hughes, K.A., Cowan, D.A. & Wilmotte, A., 2015, 'Protection of Antarctic microbial communities - "Out of sight, out of mind"', Frontiers in Microbiology 6, 151. https://doi.org/10.3389/fmicb.2015.00151 [ Links ]
Hughes, K.A. & Pertierra, L.R., 2016, 'Evaluation of non-native species policy development and implementation within the Antarctic Treaty area', Biological Conservation 200, 149-159. https://doi.org/10.1016/j.biocon.2016.03.011 [ Links ]
Hugo, E.A., McGeoch, M.A., Marshall, D.J. & Chown, S.L., 2004, 'Fine scale variation in microarthropod communities inhabiting the keystone species Azorella selago on Marion Island', Polar Biology 27(8), 466-473. https://doi.org/10.1007/s00300-004-0614-4 [ Links ]
Huiskes, A.H.L., Gremmen, N.J.M., Bergstrom, D.M., Frenot, Y., Hughes, K.A., Imura, S. et al., 2014, 'Aliens in Antarctica: Assessing transfer of plant propagules by human visitors to reduce invasion risk', Biological Conservation 171, 278-284. https://doi.org/10.1016/j.biocon.2014.01.038 [ Links ]
Hulme, P.E., 2009, 'Trade, transport and trouble: Managing invasive species pathways in an era of globalization', Journal of Applied Ecology 46(1), 10-18. https://doi.org/10.1111/j.1365-2664.2008.01600.x [ Links ]
Huntley, B.J., 1971, 'Vegetation', in E.M. Van Zinderen Bakker, J.M. Winterbottom & R.A. Dyer (eds.), Marion and Prince Edward Islands. Report on the South African biological & geological expedition 1965-1966, pp. 98-160, A.A. Balkema, Cape Town. [ Links ]
Huyser, O., Ryan, P.G. & Cooper, J., 2000, 'Changes in population size, habitat use and breeding biology of lesser sheatbills Chionis minor at Marion Island: Impacts of cats, mice and climate change?', Biological Conservation 92(3), 299-310. https://doi.org/10.1016/S0006-3207(99)00096-8 [ Links ]
Janion, C., Leinaas, H.P., Terblanche, J.S. & Chown, S.L., 2010, 'Trait means and reaction norms: The consequences of climate change/invasion interactions at the organism level', Evolutionary Ecology 24(6), 1365-1380. https://doi.org/10.1007/s10682-010-9405-2 [ Links ]
Jones, M.G.W. & Ryan, P.G., 2010, 'Evidence of mouse attacks on albatross chicks on sub-Antarctic Marion Island', Antarctic Science 22(1), 39-42. https://doi.org/10.1017/S0954102009990459 [ Links ]
Keller, R.P. & Kumschick, S., 2017, Promise and challenges of risk assessment as an approach for preventing the arrival of harmful alien species. Bothalia 47(2), a2136. https://doi.org/10.4102/abc.v47i2.2136 [ Links ]
Kloppers, F.J. & Smith, V.R., 1998, 'First Report of Botryotinia fuckeliana on Kerguelen Cabbage on the Sub-Antarctic Marion Island', Plant Disease 82(6), 710. https://doi.org/10.1094/PDIS.1998.82.6.710A [ Links ]
La Grange, J.J., 1954, 'The South African station on Marion Island', Polar Record 7(48), 155-158. https://doi.org/10.1017/S0032247400043606 [ Links ]
le Roux, P.C. & McGeoch, M.A., 2008, 'Changes in climate extremes, variability and signature on sub-Antarctic Marion Island', Climatic Change 86(3-4), 309-329. https://doi.org/10.1007/s10584-007-9259-y [ Links ]
le Roux, P.C., Ramaswiela, T., Kalwij, J.M., Shaw, J.D., Ryan, P.G., Treasure, A.M. et al., 2013, 'Human activities, propagule pressure and alien plants in the sub-Antarctic: Tests of generalities and evidence in support of management', Biological Conservation 161, 18-27. https://doi.org/10.1016/j.biocon.2013.02.005 [ Links ]
Lee, J.E. & Chown, S.L., 2007, 'Mytilus on the move: Transport of an invasive bivalve to the Antarctic', Marine Ecology Progress Series 339, 307-310. https://doi.org/10.3354/meps339307 [ Links ]
Lee, J.E. & Chown, S.L., 2009a, 'Breaching the dispersal barrier to invasion: Quantification and management', Ecological Applications 19(7), 1944-1959. https://doi.org/10.1890/08-2157.1 [ Links ]
Lee, J.E. & Chown, S.L., 2009b, 'Quantifying the propagule load associated with the construction of an Antarctic research station', Antarctic Science 21(05), 471-475. https://doi.org/10.1017/S0954102009990162 [ Links ]
Lee, J.E. & Chown, S.L., 2011, 'Quantification of intra-regional propagule movements in the Antarctic', Antarctic Science 23(04), 337-342. https://doi.org/10.1017/S0954102011000198 [ Links ]
Lee, J.E. & Chown, S.L., 2016, 'Range expansion and increasing impact of the introduced wasp Aphidius matricariae Haliday on sub-Antarctic Marion Island', Biological Invasions 18(5), 1235-1246. https://doi.org/10.1007/s10530-015-0967-3 [ Links ]
Lee, J.E., Janion, C., Marais, E., Jansen van Vuuren, B. & Chown, S.L., 2009, 'Physiological tolerances account for range limits and abundance structure in an invasive slug', Proceedings of the Royal Society B-Biological Sciences 276(1661), 1459-1468. https://doi.org/10.1098/rspb.2008.1240 [ Links ]
Lee, J.E., Slabber, S., Jansen van Vuuren, B., van Noort, S. & Chown, S.L., 2007, 'Colonisation of sub-Antarctic Marion Island by a non-indigenous aphid parasitoid Aphidius matricariae (Hymenoptera, Braconidae)', Polar Biology 30(9), 1195-1201. https://doi.org/10.1007/s00300-007-0277-z [ Links ]
Lee, J.E., Terauds, A. & Chown, S.L., 2014, 'Natural dispersal to sub-Antarctic Marion Island of two arthropod species', Polar Biology 37(6), 781-787. https://doi.org/10.1007/s00300-014-1479-9 [ Links ]
Levine, J.M., Vilà, M., Antonio, C.M.D., Dukes, J.S., Grigulis, K. & Lavorel, S., 2003, 'Mechanisms underlying the impacts of exotic plant invasions', Proceedings of the Royal Society of London. Series B: Biological Sciences 270(1517), 775-781. https://doi.org/10.1098/rspb.2003.2327 [ Links ]
Lutjeharms, J.R.E. & Ansorge, I.J., 2008, 'Oceanographic setting of the Prince Edward Islands', in S.L. Chown & P.W. Froneman (eds.), The Prince Edward Islands, pp. 17-38, SUN PReSS, Stellenbosch. [ Links ]
McClelland, G.T.W., Altwegg, R., van Aarde, R.J., Ferreira, S., Burger, A.E. & Chown, S.L., in review, 'Climate change leads to increasing population density and impacts of a key island invader'. [ Links ]
McGeoch, M.A., Butchart, S.H.M., Spear, D., Marais, E., Kleynhans, E.J., Symes, A. et al., 2010, 'Global indicators of biological invasion: Species numbers, biodiversity impact and policy responses', Diversity and Distributions 16(1), 95-108. https://doi.org/10.1111/j.1472-4642.2009.00633.x [ Links ]
McGeoch, M.A., Shaw, J.D., Terauds, A., Lee, J.E. & Chown, S.L., 2015, 'Monitoring biological invasion across the broader Antarctic: A baseline and indicator framework', Global Environmental Change 32, 108-125. https://doi.org/10.1016/j.gloenvcha.2014.12.012 [ Links ]
Ordonez, A., Wright, I.J. & Olff, H., 2010, 'Functional differences between native and alien species: A global-scale comparison', Functional Ecology 24(6), 1353-1361. https://doi.org/10.1111/j.1365-2435.2010.01739.x [ Links ]
Øvstedal, D.O. & Gremmen, N.J.M., 2011, 'The lichens of Marion and Prince Edward islands', South African Journal of Botany 67(4), 552-572. https://doi.org/10.1016/S0254-6299(15)31187-X [ Links ]
Pammenter, N.W., Drennan, P.M. & Smith, V.R., 1986, 'Physiological and anatomical aspects of photosynthesis of two Agrostis species at a sub-Antarctic island', New Phytologist 102(1), 143-160. https://doi.org/10.1111/j.1469-8137.1986.tb00806.x [ Links ]
Phiri, E.E., McGeoch, M.A. & Chown, S.L., 2009, 'Spatial variation in structural damage to a keystone plant species in the sub-Antarctic: Interactions between Azorella selago and invasive house mice', Antarctic Science 21(3), 189-196. https://doi.org/10.1017/S0954102008001569 [ Links ]
Pyšek, P. & Richardson, D.M., 2008, 'Invasive plants', in S.E. Jørgensen & B.D. Fath (eds.), Encyclopedia of ecology, Elsevier, Oxford. [ Links ]
Pyšek, P., Richardson, D.M., Pergl, J., Jarošík, V., Sixtová, Z. & Weber, E., 2008, 'Geographical and taxonomic biases in invasion ecology', Trends in Ecology & Evolution 23(5), 237-244. https://doi.org/10.1016/j.tree.2008.02.002 [ Links ]
Raymond, B., McInnes, J., Dambacher, J.M., Way, S. & Bergstrom, D.M., 2011, 'Qualitative modelling of invasive species eradication on subantarctic Macquarie Island', Journal of Applied Ecology 48(1), 181-191. https://doi.org/10.1111/j.1365-2664.2010.01916.x [ Links ]
Reaser, J.K., Meyerson, L.A., Cronk, Q., De Poorter, M., Eldrege, L.G., Green, E. et al., 2007, 'Ecological and socioeconomic impacts of invasive alien species in island ecosystems', Environmental Conservation 34(2), 98-111. https://doi.org/10.1017/S0376892907003815 [ Links ]
Richardson, D.M., Pyšek, P., Rejmánek, M., Barbour, M.G., Panetta, F.D. & West, C.J., 2000, 'Naturalization and invasion of alien plants: Concepts and definitions', Diversity and Distributions 6(2), 93-107. https://doi.org/10.1046/j.1472-4642.2000.00083.x [ Links ]
Ryan, P.G., Smith, V.R. & Gremmen, N.J.M., 2003, 'The distribution and spread of alien vascular plants on Prince Edward Island', African Journal of Marine Science 25(1), 555-562. https://doi.org/10.2989/18142320309504045 [ Links ]
Sanyika, T.W., Stafford, W. & Cowan, D.A., 2012, 'The soil and plant determinants of community structures of the dominant actinobacteria in Marion Island terrestrial habitats, Sub-Antarctica', Polar Biology 35(8), 1129-1141. https://doi.org/10.1007/s00300-012-1160-0 [ Links ]
Sax, D.F. & Gaines, S.D., 2008, 'Species invasions and extinction: The future of native biodiversity on islands', Proceedings of the National Academy of Sciences of the United States of America 105(Supplement 1), 11490-11497. https://doi.org/10.1073/pnas.0802290105 [ Links ]
Shaw, J.D., 2013, 'Southern Ocean Islands invaded: Conserving biodiversity in the world's last wilderness', in L.C. Foxcroft, P. Pyšek, D.M. Richardson & P. Genovesi (eds.), Plant invasions in protected areas: Patterns, problems and challenges, pp. 449-470, Springer Science+Business Media, Dordrecht. [ Links ]
Shaw, J.D., Spear, D., Greve, M. & Chown, S.L., 2010, 'Taxonomic homogenization and differentiation across Southern Ocean Islands differ among insects and vascular plants', Journal of Biogeography 37(2), 217-228. https://doi.org/10.1111/j.1365-2699.2009.02204.x [ Links ]
Simberloff, D., Martin, J.-L., Genovesi, P., Maris, V., Wardle, D.A., Aronson, J. et al., 2013, 'Impacts of biological invasions: What's what and the way forward', Trends in Ecology & Evolution 28(1), 58-66. https://doi.org/10.1016/j.tree.2012.07.013 [ Links ]
Slabber, S. & Chown, S.L., 2002, 'The first record of a terrestrial crustacean, Porcellio scaber (Isopoda, Porcellionidae), from sub-Antarctic Marion Island', Polar Biology 25(11), 855-858. https://doi.org/10.1016/j.jinsphys.2006.10.010 [ Links ]
Slabber, S., Worland, M.R., Leinaas, H.P. & Chown, S.L., 2007, 'Acclimation effects on thermal tolerances of springtails from sub-Antarctic Marion Island: Indigenous and invasive species', Journal of Insect Physiology 53(2), 113-125. https://doi.org/10.1016/j.jinsphys.2006.10.010 [ Links ]
Smith, V., Avenant, N. & Chown, S., 2002, 'The diet and impact of house mice on a sub-Antarctic island', Polar Biology 25(9), 703-715. [ Links ]
Smith, V.R., 1978, 'Animal-plant-soil nutrient relationships on Marion Island (Subantarctic)', Oecologia 32(2), 239-253. https://doi.org/10.1007/BF00366075 [ Links ]
Smith, V.R., 1992, 'Terrestrial slug recorded from sub-Antarctic Marion Island', Journal of Molluscan Studies 58(1), 80-81. https://doi.org/10.1093/mollus/58.1.80 [ Links ]
Smith, V.R., 2007, 'Introduced slugs and indigenous caterpillars as facilitators of carbon and nutrient mineralisation on a sub-Antarctic island', Soil Biology & Biochemistry 39(2), 709-713. https://doi.org/10.1016/j.soilbio.2006.09.026 [ Links ]
Smith, V.R., 2008, 'Energy flow and nutrient cycling in the Marion Island terrestrial ecosystem: 30 years on', Polar Record 44(3), 211-226. https://doi.org/10.1017/S0032247407007218 [ Links ]
Smith, V.R. & Steenkamp, M., 1990, 'Climatic change and its ecological implications at a subantarctic island', Oecologia 85(1), 14-24. https://doi.org/10.1007/BF00317338 [ Links ]
Springer, K., 2016, 'Methodology and challenges of a complex multi-species eradication in the sub-Antarctic and immediate effects of invasive species removal', New Zealand Journal of Ecology 40(2), 273-278. https://doi.org/10.20417/nzjecol.40.30 [ Links ]
Tecco, P.A., Díaz, S., Cabido, M. & Urcelay, C., 2010, 'Functional traits of alien plants across contrasting climatic and land-use regimes: Do aliens join the locals or try harder than them?', Journal of Ecology 98(1), 17-27. https://doi.org/10.1111/j.1365-2745.2009.01592.x [ Links ]
Treasure, A.M. & Chown, S.L., 2013, 'Contingent absences account for range limits but not the local abundance structure of an invasive springtail', Ecography 36(2), 146-156. https://doi.org/10.1111/j.1600-0587.2012.07458.x [ Links ]
Treasure, A.M. & Chown, S.L., 2014, 'Antagonistic effects of biological invasion and temperature change on body size of island ectotherms', Diversity and Distributions 20(2), 202-213. https://doi.org/10.1111/ddi.12153 [ Links ]
van Aarde, R.J., 1980, 'The diet and feeding-behavior of feral cats, Felis catus at Marion Island', South African Journal of Wildlife Research 10(3&4), 123-128. [ Links ]
van Aarde, R.J., 1986, 'A case study of an alien predator (Felis catus) introduced on Marion Island: Selective advantages', South African Journal of Antarctic Research 16(3), 113-114. [ Links ]
van Aarde, R.J., Ferreira, S.M. & Wassenaar, T.D., 2004, 'Do feral house mice have an impact on invertebrate communities on sub-Antarctic Marion Island?', Austral Ecology 29(2), 215-224. https://doi.org/10.1111/j.1442-9993.2004.01341.x [ Links ]
Van de Vijver, B., Gremmen, N. & Smith, V., 2008, 'Diatom communities from the sub-Antarctic Prince Edward Islands: Diversity and distribution patterns', Polar Biology 31(7), 795-808. https://doi.org/10.1007/s00300-008-0418-z [ Links ]
van Kleunen, M., Weber, E. & Fischer, M., 2010, 'A meta-analysis of trait differences between invasive and non-invasive plant species', Ecology Letters 13(2), 235-245. https://doi.org/10.1111/j.1461-0248.2009.01418.x [ Links ]
van Rensburg, P.J.J., 1983, 'The feeding ecology of a decreasing feral house cat, Felis catus, population at Marion Island', in W.R. Siegfried, P.R. Condy & R.M. Laws (eds.), Antarctic nutrient cycles and food webs, pp. 620-624, Springer Verlag, Berlin. [ Links ]
van Staden, W., 2011, 'Limnoecology of the freshwater algal genera (excluding diatoms) on Marion Island (sub-Antarctic)', MSc thesis, North-West University. [ Links ]
Vilà, M., Espinar, J.L., Hejda, M., Hulme, P.E., Jarošík, V., Maron, J.L. et al., 2011, 'Ecological impacts of invasive alien plants: A meta-analysis of their effects on species, communities and ecosystems', Ecology Letters 14(7), 702-708. https://doi.org/10.1111/j.1461-0248.2011.01628.x [ Links ]
Vitousek, P.M., 1988, 'Diversity and biological invasions of oceanic islands', in E.O. Wilson & F.M. Peters (eds.), Biodiversity, pp. 181-190, National Academies Press, Washington, DC. [ Links ]
Wanless, R.M., Angel, A., Cuthbert, R.J., Hilton, G.M. & Ryan, P.G., 2007, 'Can predation by invasive mice drive seabird extinctions?', Biology Letters 3(3), 241-244. https://doi.org/10.1098/rsbl.2007.0120 [ Links ]
 Correspondence:
Correspondence:
Michelle Greve
michelle_greve@yahoo.com
Received: 01 Aug. 2016
Accepted: 05 Dec. 2016
Published: 31 Mar. 2017
Note: This paper was initially delivered at the 43rd Annual Research Symposium on the Management of Biological Invasions in South Africa, Goudini Spa, Western Cape, South Africa on 18-20 May 2016.
Appendix 1
Terrestrial invasions on Sub-Antarctic Marion and Prince Edward Islands M. Greve, R. Mathakutha, C. Steyn & S.L. Chown





References
Blackburn, T.M., Essl, F., Evans, T., Hulme, P.E., Jeschke, J.M. et al., 2014, 'A unified classification of alien species based on the magnitude of their environmental impacts', PLoS Biology, 12(5), e1001850. [ Links ]
Blackburn, T.M., Pyšek, P., Bacher, S., Carlton, J.T., Duncan, R.P., Jarošík, V. et al., 2011, 'A proposed unified framework for biological invasions', Trends in Ecology & Evolution, 26(7), 333-339. [ Links ]


{kind=link}
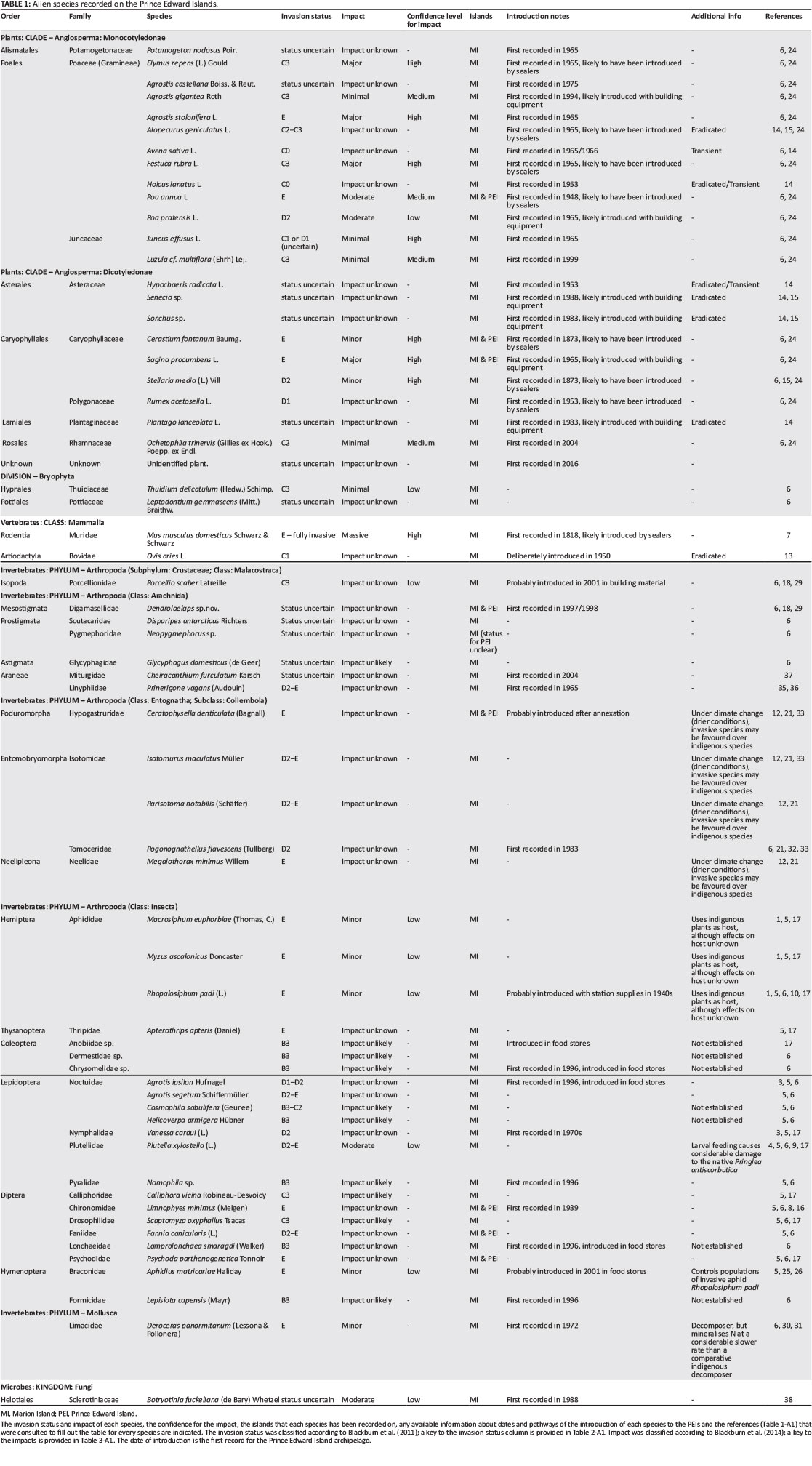{kind=link}
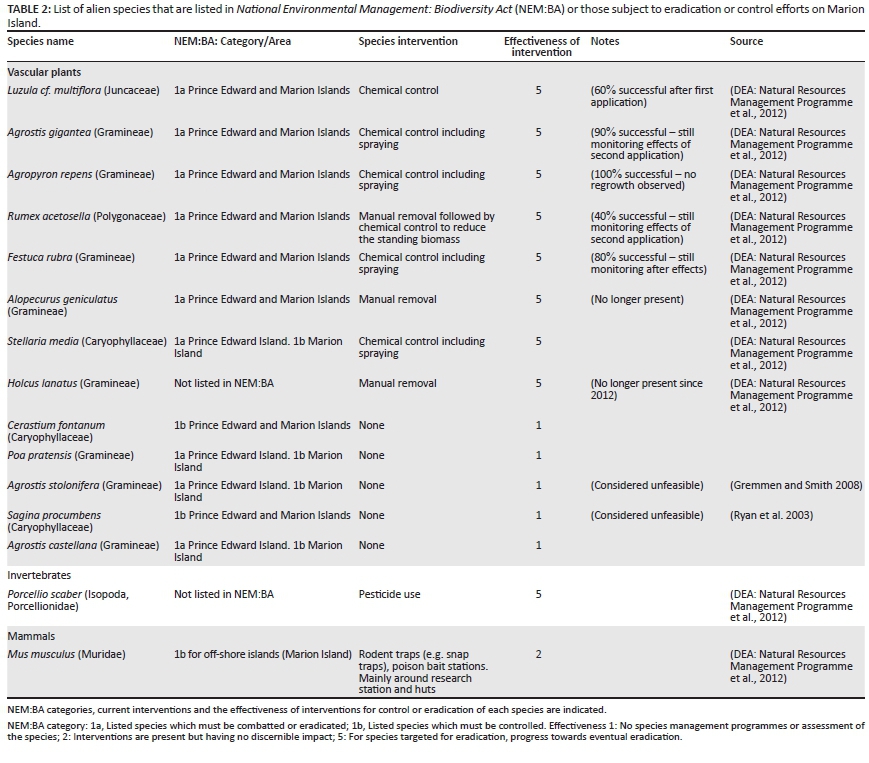{kind=link}