










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.47 n.2 Pretoria 2017
http://dx.doi.org/10.4102/abc.v47i2.2157
ORIGINAL RESEARCH
The balance of trade in alien species between South Africa and the rest of Africa
Katelyn T. FaulknerI, II; Brett P. HurleyIII, IV; Mark P. RobertsonII; Mathieu RougetV; John R.U. WilsonI, VI
IInvasive Species Programme, South African National Biodiversity Institute, Kirstenbosch Research Centre, South Africa
IICentre for Invasion Biology, Department of Zoology and Entomology, University of Pretoria, South Africa
IIIForestry and Agricultural Biotechnology Institute (FABI), University of Pretoria, South Africa
IVDepartment of Zoology and Entomology, University of Pretoria, South Africa
VCentre for Invasion Biology, School of Agricultural, Earth and Environmental Sciences, University of KwaZulu-Natal, South Africa
VICentre for Invasion Biology, Department of Botany and Zoology, Stellenbosch University, South Africa
ABSTRACT
BACKGROUND: Alien organisms are not only introduced from one biogeographical region to another but also spread within regions. As South Africa shares land borders with six countries, multiple opportunities exist for the transfer of alien species between South Africa and other African countries; however, the direction and importance of intra-regional spread is unclear.
OBJECTIVES: The aim of this study was to gain a greater understanding of the introduction of alien species into Africa and the spread of species between South Africa and other African countries.
METHOD: We developed scenarios that describe the routes by which alien species are introduced to and spread within Africa and present case studies for each. Using data from literature sources and databases, the relative importance of each scenario for alien birds and insect pests of eucalypts was determined, and the direction and importance of intra-regional spread was assessed.
RESULTS: Alien species from many taxonomic groups have, through various routes, been introduced to and spread within Africa. For birds and eucalypt insect pests, the number of species spreading in the region has recently increased, with South Africa being a major recipient of birds (14 species received and 5 donated) and a major donor of eucalypt insect pests (1 species received and 10 donated). For both groups, many introduced species have not yet spread in the region.
CONCLUSION: The intra-regional spread of alien species in Africa represents an important and possibly increasing threat to biosecurity. To address this threat, we propose a framework that details how African countries could cooperate and develop a coordinated response to alien species introductions.
Introduction
The movement of goods and people around the world is facilitating the introduction of organisms to regions where they are not native. Although many alien organisms are introduced directly from one biogeographical region to another ('inter-regional introduction'), the spread of species within biogeographical regions also contributes to biological invasions ('intra-regional spread') (Chiron, Shirley & Kark 2010; Hurley et al. 2016; Jaksic et al. 2002; Roques et al. 2016 in this article, the biogeographical region of interest is continental Africa). Relatively high propagule pressure [i.e. the number of individuals introduced and the number of introduction events for a specific species (Lockwood, Cassey & Blackburn 2005)] and short geographical distances mean that once an organism has been introduced to a region, further natural or human-aided spread is likely (Garnas et al. 2016; Hurley et al. 2016; Jaksic et al. 2002; Roques et al. 2016). Furthermore, organisms that are native to a biogeographical region might spread within the region, either naturally or with the aid of humans, to areas where they are not native (Chiron et al. 2010).
The intra-regional spread of species is often asymmetrical [i.e. one country donates more species than it receives (Ferus et al. 2015; Jaksic et al. 2002)] and, under some circumstances, introductions through intra-regional spread may be more common than those that occur through inter-regional introduction (Chiron et al. 2010). However, such patterns are the result of historical economic and socio-political processes and so can vary over time (Chiron et al. 2010; Essl et al. 2011; Roques et al. 2016). Furthermore, as properties linked with invasion success (e.g. likelihood of enemy release and propagule pressure) vary across dispersal pathways (e.g. extreme long-distance or leading-edge dispersal; see Wilson et al. 2009), whether an organism is introduced through inter-regional introduction or intra-regional spread can have consequences for its invasion success. To develop and improve efforts aimed at preventing or mitigating the introduction of invasive species, it is therefore important to identify the types of introduction within a region and determine their relative importance and direction [also see the Convention on Biological Diversity's Aichi Target 9 (UNEP 2011)].
Many organisms have been directly introduced to South Africa from other continents [e.g. the Sirex woodwasp (Sirex noctilio), which is native to Eurasia and northern Africa, was introduced to South Africa from Oceania and South America (Boissin et al. 2012), and the harlequin ladybird (Harmonia axyridis), which is native to Asia, was introduced to South Africa from North America (Lombaert et al. 2010)]. Given that South Africa shares land borders with six other African countries, multiple opportunities exist for species to spread between South Africa and other African countries, either through natural dispersal or with the aid of various human-related transport vectors (e.g. air, sea and land transport vectors). However, the relative importance of inter-regional introduction and intra-regional spread is currently not clear, and whether South Africa is primarily a donor or recipient of alien species is also unknown.
In an effort to gain a greater understanding of the movements of alien species into and within Africa, we aimed to (1) develop introduction route scenarios that describe how alien species might have been introduced to the region and spread between South Africa and elsewhere in Africa; (2) demonstrate these scenarios using case studies; (3) use the scenarios to quantify, for selected groups, the importance and direction of intra-regional spread; (4) determine if these patterns have changed through time; and (5) propose a framework for trans-boundary collaboration in biosecurity that could address the threat posed to Africa by the intra-regional spread of alien species.
Research method and design
Introduction route scenarios
Six introduction route scenarios that describe how alien species might have been introduced to the region and spread between continental South Africa (SA) and elsewhere in continental Africa (AF) were developed and examples identified. In these scenarios, both natural dispersal and the human-aided movement of species within the continent were considered as intra-regional spread. Furthermore, although introduction and spread through all human-related transport vectors were considered (i.e. land, air and sea transport vectors), these vectors were not discriminated in the scenarios. The scenarios describe introduction routes where species have been introduced to SA or AF from other regions, and consider whether subsequent intra-regional spread between SA and AF occurred. Also described are instances where species that are native to either SA or AF have spread between the subregions to areas where they are not native. Details on the introduction route scenarios and examples of species for each scenario are shown in Figure 1, and the details for each example are provided in Appendix 2, Figure 1-A2 to Figure 6-A2.
Importance of introductions into Africa versus spread within Africa
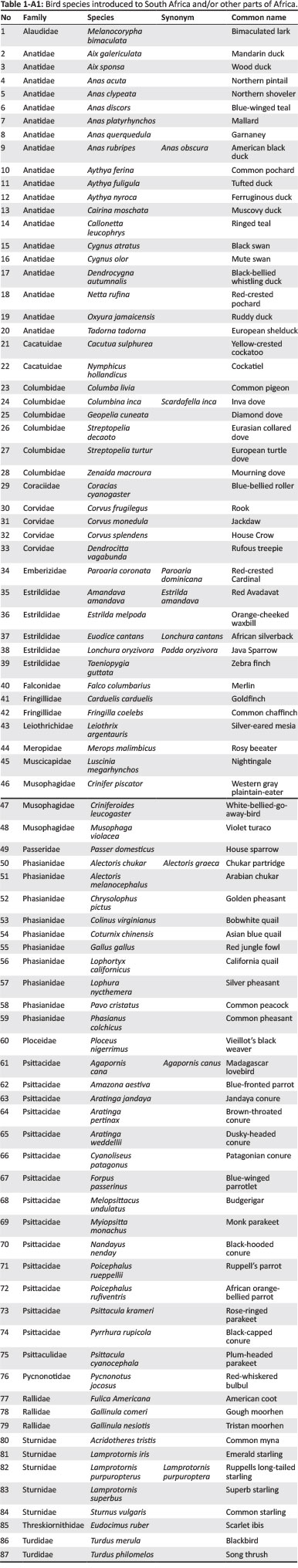
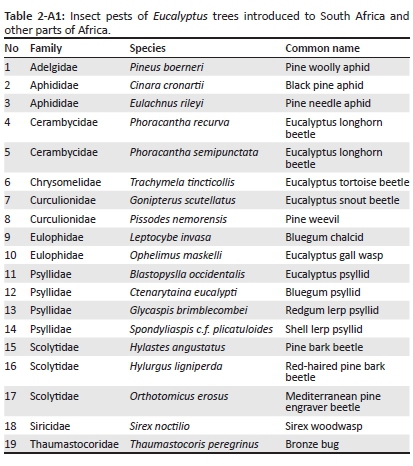
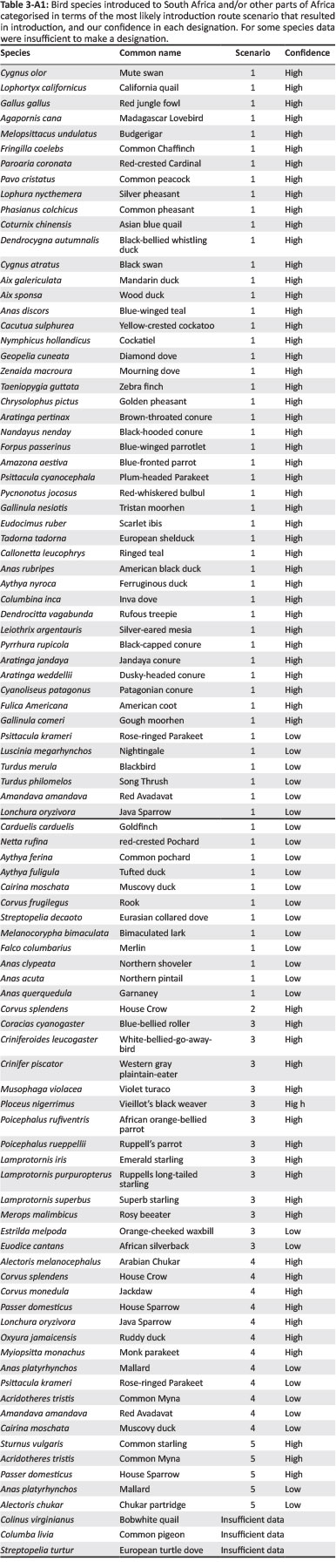
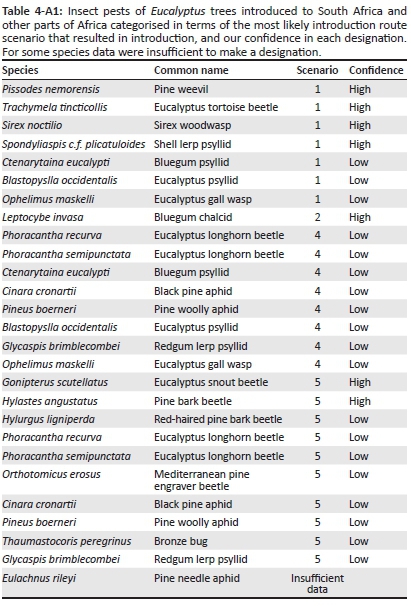
For alien species in SA and AF, information on their native and introduced range (in Africa and elsewhere in the world), descriptions of introduction and spread as well as introduction data (introduction source, number of introductions, pathway of introduction and date of introduction or first record) were used to categorise species in terms of the most likely introduction route scenario that resulted in introduction. As detailed species-level introduction data are often lacking (Faulkner et al. 2015), we focused on two groups for which these data could be obtained: birds and insect pests of Eucalyptus trees (see Tables 1-A1 and 2-A1 for species lists). For birds, data were extracted from South African (i.e. Dean 2000; Peacock, van Rensburg & Robertson 2007; Picker & Griffiths 2011; van Rensburg et al. 2011) and global sources (i.e. CAB International 2016; Lever 1987, 2005; Long 1981). For each eucalypt insect pest in South Africa, date of first record for southern African countries was obtained from Bush et al. (2016), Wingfield et al. (2008) and local authorities, and other data were extracted from South African (i.e. Picker & Griffiths 2011) and global information sources (i.e. CAB International 2016).
Re-introductions [e.g. helmeted guineafowl (Numida meleagris) in South Africa (Lever 1987; Long 1981)] and extralimital populations [species that have been translocated within the subregion where they are native to parts of that subregion where they are not native; e.g. red-eyed dove (Streptopelia semitorquata) in South Africa (Lever 1987; Long 1981)] were not included. For species that have been introduced to the region multiple times, all applicable scenarios were recorded (and therefore the total count of species for the scenarios can be larger than the total number of species investigated). Furthermore, this means that although a species might not have spread between the two subregions, as a result of independent introductions it might still occur in both SA and AF. For some introductions, it was clear which scenario was applicable [e.g. scenario 5 is clearly applicable for the common starling (Sturnus vulgaris) which was introduced in the 1800s from the United Kingdom to SA and then spread into neighbouring countries, i.e. AF], but for others this was not the case [e.g. the Indian subspecies of the rose-ringed parakeet (Psittacula krameri) was introduced to SA in the 1800s and then later to Egypt (i.e. AF), it is likely that these introductions were independent (i.e. both scenarios 1 and 4 are applicable); however, it is possible that the birds in Egypt came from SA (i.e. scenario 5)]. In an effort to account for this uncertainty, we categorised each introduction according to the most likely scenario (i.e. scenario 5 for S. vulgaris and scenarios 1 and 4 for P. krameri) and then rated our confidence in each designation as high or low. A high confidence rating was assigned when the scenario was clear (e.g. for S. vulgaris), and low confidence was assigned when more than one scenario was possible (e.g. for P. krameri). For some species, there were insufficient data to make a designation, for example, the common pigeon (Columba livia) is native to parts of North Africa and has also been introduced widely on the continent; however, details are imprecise for many of these introductions and thus multiple, equally likely scenarios are possible (Lever 2005). These species were recorded as having insufficient data, and they were not assigned an introduction route scenario.
While some of the scenarios involve only inter-regional introductions or intra-regional spread, others involve combinations of the two introduction types (i.e. a species is introduced to AF or SA and also spreads between SA and AF; see Figure 3). Therefore, the results for a number of the scenarios had to be combined to determine the relative importance of the two types of introduction. For instance, to determine the total number of species introduced from another biogeographical region to SA, results for scenarios 1 and 5 had to be combined (Figure 3). However, as the six scenarios provide useful details that are lost when combined (e.g. whether the species is native or alien to the region), we evaluated alien species movements in terms of both the scenarios and the types of introduction.

A generalised linear model (Poisson error distribution and log link) was used to analyse a two-way contingency table of species counts (Crawley 2007) and test the association between organism type (birds and eucalypt insect pests) and scenario. The relative importance of inter-regional introduction and intra-regional spread between SA and AF was assessed by calculating the number of bird and eucalypt insect pest species directly introduced from another region to SA (sum of the counts for scenarios 1 and 5) and AF (sum of the counts for scenarios 2 and 4), and the number of species that spread into SA from AF (sum of the counts for scenarios 2 and 3) and vice versa (sum of the counts for scenarios 5 and 6). A generalised linear model (Poisson error distribution and log link) was used to analyse a three-way contingency table of species counts and test the association between organism type (birds and eucalypt insect pests), recipient subregion (SA and AF) and introduction type (inter-regional introduction and intra-regional spread). To evaluate whether the relative importance of inter-regional introduction and intra-regional spread has varied over time, date of introduction or first record data were used to designate introductions into 50-year time periods, and for each period, the number of birds and eucalypt insect pests introduced to SA and AF through inter-regional introduction and intra-regional spread was determined. All generalised linear models were checked for overdispersion (Crawley 2007; Zuur et al. 2009), but no instances were noted. Counts that were significantly different from what might be expected based on chance alone were identified by calculating the standardised adjusted residuals and comparing these values to the critical values of the normal distribution (Bewick, Cheek & Ball 2004; Everitt 1977). In an effort to determine the influence of uncertainty on results, analyses were performed twice: using all the data and using a subset with only designations made with high certainty.
Results
Importance of introductions into Africa versus spread within Africa
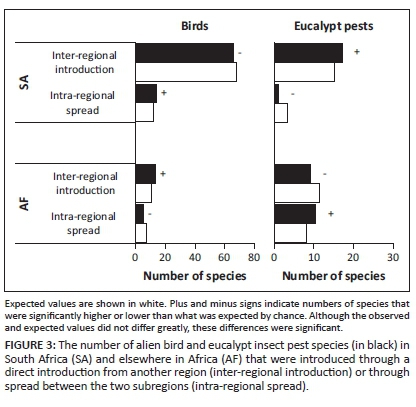
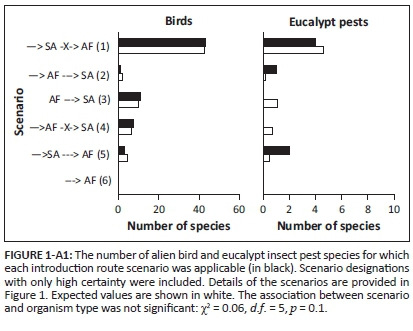
Birds and eucalypt insect pests have been introduced through various introduction routes (Tables 3-A1 and 4-A1), but different scenarios were common for the two groups (significant association between scenario and organism type: χ2 = 30.6, d.f. = 5, p < 0.001). Many bird species that have been introduced to SA from another biogeographical region did not spread to AF (scenario 1, see Figure 2); however, four of these species have been independently introduced to AF (scenario 4). While the number of bird species for which scenario 1 was applicable was significantly higher than expected by chance, so too was the number of bird species that are native to AF that spread into SA (scenario 3, see Figure 2). For eucalypt insect pests, most species, and a significantly higher number than expected, were introduced to SA from another region and then subsequently spread into AF (scenario 5, see Figure 2). A significantly higher number of eucalypt insect pests than expected by chance were also directly introduced to AF from another region and then did not spread into SA (scenario 4, see Figure 2). However, all of these species have also been introduced to SA (scenario 1). Multiple scenarios were applicable for eight bird and eight eucalypt insect pest species. There were few instances of insufficient data (3% for birds and 4% for eucalypt insect pests, see Figure 2). For birds, 68% of the scenario designations were made with high certainty, but for eucalypt insect pests, this was only the case for 26% of designations. Consequently, the results of the statistical analysis differed when only scenario designations with high certainty were included (the association between scenario and organism type was no longer significant), but for birds, the identified pattern (i.e. scenario 1 dominated, but for many species scenario 3 was applicable) remained the same (Figure 1-A1).
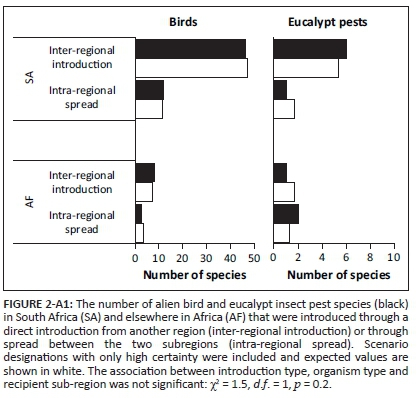
The relative importance of inter-regional introduction and intra-regional spread differed for birds and eucalypt insect pests and also varied based on the recipient subregion (Figure 3; significant association between introduction type, organism type and recipient subregion: χ2 = 4.3, d.f. = 1, p = 0.04). Based on the species for which the date of introduction data were available (75% for birds and 89% for eucalypt insect pests), it appears that for alien birds and eucalypt insect pests in SA and AF, the relative importance of inter-regional introduction and intra-regional spread changed over time (Figure 4). Although most alien bird species in SA were introduced from other regions, this number was significantly lower than expected by chance, and the number that spread from AF into SA was significantly higher than expected (Figure 3). Additionally, since 2000, more species have spread from AF to SA than have been introduced from other regions (Figure 4). For eucalypt insect pests in SA, a significantly higher number than expected have been introduced from other regions (Figure 3), but since 2000, one species has also spread from AF to SA (Figure 4). A significantly higher number of bird species were introduced from other regions to AF than was expected (Figure 3), with this number being higher, for all time periods since 1900, than the number that have spread from SA into AF (Figure 4). The number of eucalypt insect pests that spread from SA to AF was significantly higher than that expected by chance (Figure 3), with more species spreading from SA to AF since 2000 than the number introduced from other regions (Figure 4). In the spread of alien species between SA and AF, SA was the major recipient of birds but the major donor of eucalypt insect pests (Figure 3). When only designations with high certainty were included in the analysis, the statistical results changed (the association between introduction type, organism type and recipient subregion was no longer significant), but the identified patterns were the same (i.e. most birds and eucalypt insect pests in SA and birds in AF were inter-regional introductions, but most eucalypt insect pests in AF spread in from SA, and SA was a major recipient of birds but a major donor of eucalypt insect pests; Figure 2-A1).
Discussion
The patterns of movement of alien species are often complex (Hurley et al. 2016) and, in line with this, our case studies and results show that alien organisms in Africa have been introduced through various introduction routes (e.g. see Measey et al. 2017 for a discussion on amphibians; and Visser et al. 2017 for grasses). Although many species are introduced to the continent directly from other regions, species are also spreading within Africa, with the relative importance and direction of spread varying across organisms and over time. This poses a challenge to biosecurity that needs to be addressed (see Keller & Kumschick 2017).
Importance of introductions to Africa versus spread within Africa
Many birds and eucalypt insect pests have been introduced to either SA or AF but have not yet spread between the two subregions. Similarly, many alien species in other regions [e.g. plants (Lambdon et al. 2008), birds (Chiron et al. 2010) and insects (Roques et al. 2016) in Europe] have not spread from the country where they were introduced. The spread of these species might be limited by a variety of factors, including the environment and their dispersal capabilities (Roques et al. 2016), but in many cases, it might simply be a matter of time before they spread across national boundaries. Most bird species that were introduced to SA and subsequently spread into AF were introduced over 100 years ago, while those species that have not yet spread tend to have been introduced during or after the 1970s. The future spread of these species likely represents a major invasion debt (Rouget et al. 2016).
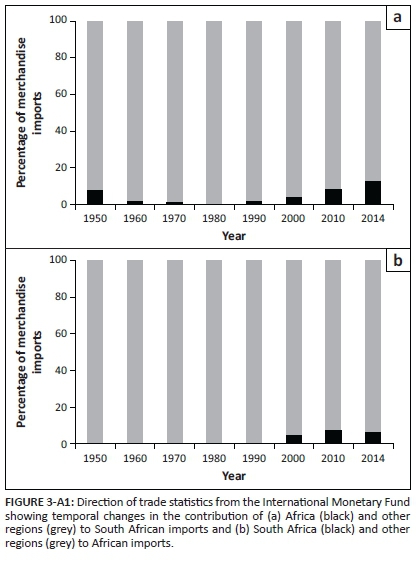
Although introductions from other regions dominated in most cases, the spread of species between SA and AF has recently increased in importance. This trend might be driven by recent growth in trade between these two subregions (Figure 3-A1). The link between socio-economic factors and the introduction and spread of alien species is well documented (Essl et al. 2011) and, for example, political and economic changes in Europe (e.g. the cold war and the later opening of borders to movement and trade) have influenced the spread of alien bird and insect species in the region (Chiron et al. 2010; Roques et al. 2016).
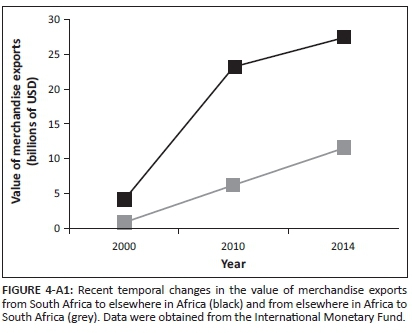
South Africa is one of a few countries that serve as major introduction points for eucalypt insect pests (Hurley et al. 2016). Additionally, South Africa currently exports more goods to other African countries than it imports (Figure 4-A1). Thus, it is not surprising that South Africa appears to be a major donor in the intra-regional spread of eucalypt insect pests. Contrary to the dominant direction of trade, South Africa is the major recipient in the intra-regional spread of birds. However, as birds are often introduced intentionally, their movement patterns might be less likely than those of eucalypt insect pests (usually introduced accidentally), to reflect coarse trends in trade (but, see Seebens et al. 2015 for predictors of global flows of naturalised plants).
The results discussed above are based on only two groups for which some historical data could be obtained. Because of data quality issues, using historical data to determine the introduction routes of alien species can lead to imprecise inferences (for information on genetic techniques, see Estoup & Guillemaud 2010). For example, countries differ with regard to their surveillance and monitoring activities (Latombe et al. in press), and as a consequence, species that have been recorded in South Africa first might not have been introduced directly from another region, but might instead have spread into the country from elsewhere in Africa where their introduction was not detected. To get an indication of how data quality impacted our results, we conducted the analysis on the full dataset and on a subset in which we had high confidence. Although the results of these analyses differed, the overall conclusions were the same (i.e. that inter-regional introductions dominate but that intra-regional spread is important, and that South Africa is a major donor for eucalypt insect pests but a major recipient for birds). Furthermore, by focusing on birds and eucalypt insect pests, we were able to highlight that organisms are being introduced to and are spreading within Africa, a pattern which, we show using our case studies, is true for a wide variety of organisms.
Intra-African spread as a biosecurity threat
Although inter-regional introductions and intra-regional spread may, in general, be increasing as a result of increased global travel and trade (Hurley et al. 2016), an increase in the number of species spreading within Africa (as shown here for birds and eucalypt insect pests) might pose a particularly high biosecurity threat. Shorter geographical distances, and higher propagule pressure and environmental similarity mean that the chances of naturalisation might be higher for species spreading within a region than for those introduced directly from other regions (van Kleunen et al. 2015). In keeping with this, for many regions (including Africa), a higher number of plant species than expected are native to a part of the region but have been introduced to and have become naturalised in other parts of the region where they are not native (van Kleunen et al. 2015). Furthermore, as many alien species present in Africa may still establish and spread (i.e. establishment and spread debt, see Rouget et al. 2016), it is likely that the biosecurity threat posed by intra-African spread will continue to increase.
The increasing number of species that have spread between South Africa and the rest of Africa highlights that current efforts in South Africa to prevent or reduce introductions from other African countries are likely insufficient. This potential weakness in South Africa's biosecurity deserves consideration in the first National Status Report on Biological Invasions, and in any future plans to manage South Africa's pathways of introduction (Wilson et al. 2017).
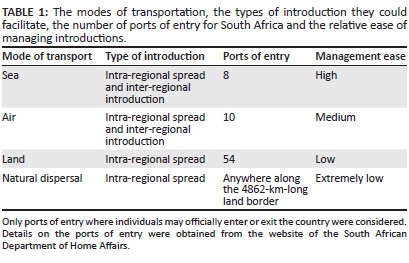
Unfortunately, preventing intra-regional spread is particularly difficult. Organisms from outside Africa can only be transported directly to South Africa by air or sea, and thus to prevent their introduction, border control only needs to be implemented at 18 official ports of entry (Table 1). In contrast, species in Africa can spread into South Africa through natural dispersal or with the aid of land, sea or air transportation (Table 1). Thus, to prevent organisms from spreading into the country, not only does border control need to be implemented at 73 official ports of entry, but South Africa's 4862-km-long land boundary (Central Intelligence Agency 2015) also needs to be managed (Table 1).
To overcome this problem, and better manage biological invasions in the region, a coordinated regional response is needed. Attempts have been made in other regions to achieve this and, for example, the Chilean and Argentinian governments have developed joint research and control programmes for alien mammals (Jaksic et al. 2002), while an early warning and response framework has been developed for Europe (Genovesi et al. 2010). Based on the European system, we have developed a framework, shown in Figure 5, to coordinate the response of African countries to alien species introductions. The framework details the activities that countries should perform before (i.e. pre-border activities) and after (i.e. post-border activities) an alien species is detected; highlights when communication and information sharing between countries is required (e.g. report the detection of an alien species); and details how countries should respond to the introduction of a potential invader based on its type of spread (i.e. unaided or intentionally or unintentionally aided by humans). To achieve these actions, various types of information (e.g. the status of a species and its invasion history) and expertise (e.g. taxonomic experts for identifications) are required. As data availability on alien species varies across countries (McGeoch et al. 2010), we recommend that the required data should be maintained in a regional information system, which is regularly updated and to which information from global databases contributes [e.g. CABI's Invasive Species Compendium and the Global Invasive Species Information Network (GISIN), also see Lucy et al. 2016]. Such a database would improve not only the availability of alien species data but also, if standards are put in place, the quality of the data. Finally, it is unlikely that all of the required expertise will be available in every African country, for example, no country will have taxonomic experts for every taxonomic group (Klopper, Smith & Chikuni 2002). Regional cooperation, particularly in the training of personnel and the exchange of experts, is therefore required. In an effort to achieve this for taxonomy, regional networks [e.g. SABONET (Willis & Huntley 2001), which came to a close in 2005] and international initiatives [e.g. the Global Taxonomy Initiative (Secretariat of the Convention on Biological Diversity 2010)] have already been established.
Conclusion
A wide variety of alien species have been introduced to Africa and have spread within the region, with the movement patterns of these species varying across organisms and over time. Although direct introductions from other regions remain a concern, the number of species spreading within the region appears to be increasing, and these species probably pose a particularly high biosecurity threat. As preventing the intra-African spread of species is at best difficult, African countries need to cooperate and coordinate their responses. Achieving this requires communication, the development and implementation of standardised methods and systems, and political will. As the efficacy of a country's biosecurity greatly influences that of its neighbours, such an endeavour would benefit all of the countries involved.
Acknowledgements
We thank Georg Goergen, Jesse Kalwij, Ingrid Minnaar and Olaf Weyl for providing photos of example species. We also thank Donald Chungu (Copperbelt University, Zambia), Tembani Mduduzi (Forest Research Centre, Zimbabwe), Peter Kiwuso (Forest Research Institute, Uganda), Gerald Meke (FRIM, Malawi) and Eston Mutitu (KEFRI, Kenya) for providing information on the detection date of eucalypt insect pests in their respective countries.
This work was supported by the South African National Department of Environment Affairs through its funding of the South African National Biodiversity Institute's Invasive Species Programme. Additional funding was provided by the DST-NRF Centre for Invasion Biology. M.R. acknowledges funding from the South African Research Chairs Initiative of the Department of Science and Technology and National Research Foundation of South Africa.
Competing interests
The authors declare that they have no financial or personal relationship(s) that may have inappropriately influenced them in writing this article.
Author(s) contributions
K.T.F., M.P.R. and J.R.U.W. developed the scenarios. K.T.F. identified the examples. K.T.F. and B.P.H. collected the data. K.T.F. analysed the data. K.T.F., B.P.H., M.P.R., M.R. and J.R.U.W. wrote the manuscript.
References
Bewick, V., Cheek, L. & Ball, J., 2004, 'Statistics review 8: Qualitative data - Tests of association', Critical Care 8(1), 46-53. https://doi.org/10.1186/cc2428 [ Links ]
Boissin, E., Hurley, B., Wingfield, M.J., Vasaitis, R., Stenlid, J., Davis, C. et al., 2012, 'Retracing the routes of introduction of invasive species: The case of the Sirex noctilio woodwasp', Molecular Ecology 21(23), 5728-5744. https://doi.org/10.1111/mec.12065 [ Links ]
Bourgeois, K., Suehs, C.M., Vidal, E. & Médail, F., 2005, 'Invasional meltdown potential: Facilitation between introduced plants and mammals on French Mediterranean islands', Écoscience 12(2), 248-256. https://doi.org/10.2980/i1195-6860-12-2-248.1 [ Links ]
Brandes, D., 2001, Urban flora of Sousse (Tunisia), Braunschweig University Library Digitale Bibliothek. [ Links ]
Bromilow, C., 2010, Problem plants and alien weeds of South Africa, 3rd edn., BRIZA Publications, Pretoria. [ Links ]
Brown, P.M.J., Thomas, C.E., Lombaert, E., Jeffries, D.L., Estoup, A. & Lawson Handley, L.-J., 2011, 'The global spread of Harmonia axyridis (Coleoptera: Coccinellidae): Distribution, dispersal and routes of invasion', BioControl 56, 623-641. https://doi.org/10.1007/s10526-011-9379-1 [ Links ]
Bush, S.J., Slippers, B., Neser, S., Harney, M., Dittrich-Schröder, G. & Hurley, B.P., 2016, 'Six recently recorded Australian insects associated with eucalyptus in South Africa', African Entomology 24(2), 539-544. https://doi.org/10.4001/003.024.0539 [ Links ]
CAB International, 2000a, Prostephanus truncatus, invasive species compendium, viewed 10 February 2016, from http://www.cabi.org/isc/datasheet/44524. [ Links ]
CAB International, 2000b, Medicago polymorpha, invasive species compendium, viewed 22 February 2016, from http://www.cabi.org/isc/datasheet/33031 [ Links ]
CAB International, 2000c, Carpobrotus edulis, invasive species compendium, viewed 17 February 2016, from http://www.cabi.org/isc/datasheet/10648 [ Links ]
CAB International, 2016, Invasive species compendium, viewed 5 January 2016, from http://www.cabi.org/isc [ Links ]
Canonico, G.C., Arthington, A., McCrary, J.K. & Thieme, M.L., 2005, 'The effects of introduced tilapias on native biodiversity', Aquatic Conservation: Marine and Freshwater Ecosystems 15, 463-483. https://doi.org/10.1002/aqc.699 [ Links ]
Carnegie, A.J., Matsuki, M., Haugen, D.A., Hurley, B.P., Ahumada, R., Klasmer, P. et al., 2006, 'Predicting the potential distribution of Sirex noctilio (Hymenoptera: Siricidae), a significant exotic pest of Pinus plantations', Annals of Forest Science 63, 119-128. https://doi.org/10.1051/forest:2005104 [ Links ]
Central Intelligence Agency, 2015, The CIA world factbook 2016, Skyhorse Publishing, New York. [ Links ]
Chiron, F., Shirley, S.M. & Kark, S., 2010, 'Behind the Iron Curtain: Socio-economic and political factors shaped exotic bird introductions into Europe', Biological Conservation 143, 351-356. https://doi.org/10.1016/j.biocon.2009.10.021 [ Links ]
Crawley, M.J., 2007, The R book, Wiley, Chichester. [ Links ]
D'Antonio, C.M., 1990, 'Seed production and dispersal in the non-native, invasive succulent Carpobrotus edulis (Aizoaceae) in coastal strand communities of central California', Journal of Applied Ecology 27(2), 693-702. https://doi.org/10.2307/2404312 [ Links ]
de Moor, I.J. & Bruton, M.N., 1988, Atlas of alien and translocated indigenous aquatic animals in southern Africa, Council for Scientific and Industrial Research, Pretoria. [ Links ]
Deacon, J., 1986, 'Human settlement in South Africa and archaeological evidence for alien plants and animals', in I.A.W. Macdonald, F.J. Kruger & A.A. Ferrar (eds.), The ecology and management of biological invasions in southern Africa, pp. 3-19, Oxford University Press, Cape Town. [ Links ]
Dean, W.R.J., 2000, 'Alien birds in southern Africa: What factors determine success?', South African Journal of Science 96, 9-14. [ Links ]
Department of Home Affairs, 2016, South African ports of entry, viewed 9 June 2016, from http://www.dha.gov.za/index.php/immigration-services/south-african-ports-of-entry. [ Links ]
Essl, F., Dullinger, S., Rabitsch, W., Hulme, P.E., Hülber, K., Jarošík, V. et al., 2011, 'Socioeconomic legacy yields an invasion debt', Proceedings of the National Academy of Sciences of the United States of America 108(1), 203-207. https://doi.org/10.1073/pnas.1011728108 [ Links ]
Estoup, A. & Guillemaud, T., 2010, 'Reconstructing routes of invasion using genetic data: Why, how and so what?', Molecular Ecology 19, 4113-4130. https://doi.org/10.1111/j.1365-294X.2010.04773.x [ Links ]
Everitt, B.S., 1977, The analysis of contingency tables, Chapman and Hall Ltd, London. [ Links ]
Falleh, H., Ksouri, R., Boulaaba, M., Guyot, S., Abdelly, C. & Magné, C., 2012, 'Phenolic nature, occurrence and polymerization degree as marker of environmental adaptation in the edible halophyte Mesembryanthemum edule', South African Journal of Botany 79, 117-124. https://doi.org/10.1016/j.sajb.2011.10.001 [ Links ]
Faulkner, K.T., Spear, D., Robertson, M.P., Rouget, M. & Wilson, J.R.U., 2015, 'An assessment of the information content of South African alien species databases', Bothalia: African Biodiversity and Conservation 45(1). https://doi.org/10.4102/abc.v45i1.1103 [ Links ]
Ferus, P., Culiţă, S., Eliáš, P., Konôpková, J., Ďurišová, L., Samuil, C. et al., 2015, 'Reciprocal contamination by invasive plants: Analysis of trade exchange between Slovakia and Romania', Biologia 70(7), 893-904. https://doi.org/10.1515/biolog-2015-0102 [ Links ]
Garnas, J.R., Auger-Rozenberg, M.-A., Roques, A., Bertelsmeier, C., Wingfield, M.J., Saccaggi, D.L. et al., 2016, 'Complex patterns of global spread in invasive insects: Eco-evolutionary and management consequences', Biological Invasions 18(4), 921-933. https://doi.org/10.1007/s10530-016-1082-9 [ Links ]
Genovesi, P., Scalera, R., Brunel, S., Roy, D. & Solarz, W., 2010, Towards an early warning and information system for invasive alien species (IAS) threatening biodiversity in Europe, European Environment Agency Technical Report, European Environmental Agency, Copenhagen. [ Links ]
Greuter, W. & Domina, G., 2015, 'Checklist of vascular plants collected during the 12th "Iter Mediterraneum" in Tunisia, 24 March-4 April 2014', Bocconea 27(1), 21-61. [ Links ]
Hodges, R.J., 1986, 'The biology and control of Prostephanus truncatus (Horn) (Coleoptera: Bostrichidae) - A destrutive storage pest with an increasing range', Journal of Stored Products Research 22(1), 1-14. https://doi.org/10.1016/0022-474X(86)90040-8 [ Links ]
Hurley, B.P., Croft, P., Verleur, M., Wingfield, M.J. & Slippers, B., 2012, 'The control of the Sirex woodwasp in diverse environments: The South African experience', in B. Slippers, P. de Groot & M.J. Wingfield (eds.), The Sirex woodwasp and its fungal symbiont: Research and management of a worldwide invasive pest, pp. 247-264, Springer, Dordrecht, The Netherlands. [ Links ]
Hurley, B.P., Garnas, J., Wingfield, M.J., Branco, M., Richardson, D.M. & Slippers, B., 2016, 'Increasing numbers and intercontinental spread of invasive insects on eucalypts', Biological Invasions 18(4), 921-933. https://doi.org/10.1007/s10530-016-1081-x [ Links ]
Hurley, B.P., Slippers, B. & Wingfield, M.J., 2007, 'A comparison of control results for the alien invasive woodwasp, Sirex noctilio, in the Southern Hemisphere', Agricultural and Forest Entomology 9, 159-171. https://doi.org/10.1111/j.1461-9563.2007.00340.x [ Links ]
Jaksic, F.M., Iriarte, J.A., Jiménez, J.E. & Martínez, D.R., 2002, 'Invaders without frontiers: Cross-border invasions of exotic mammals', Biological Invasions 4, 157-173. https://doi.org/10.1023/A:1020576709964 [ Links ]
Keller, R.P. & Kumschick, S., 2017, 'Promise and challenges of risk assessment as an approach for preventing the arrival of harmful alien species', Bothalia 47(2), a2136. https://doi.org/10.4102/abc.v47i2.2136 [ Links ]
Kenis, M., Roy, H.E., Zindel, R. & Majerus, M.E.N., 2008, 'Current and potential management strategies against Harmonia axyridis', BioControl 53(1), 235-252. https://doi.org/10.1007/s10526-007-9136-7. [ Links ]
Klopper, R.R., Smith, G.F. & Chikuni, A.C., 2002, 'The global taxonomy initiative in Africa', Taxon 51, 159-165. https://doi.org/10.2307/1554974 [ Links ]
Lambdon, P.W., Pyšek, P., Basnou, C., Hejda, M., Arianoutsou, M., Essl, F. et al., 2008, 'Alien flora of Europe: Species diversity, temporal trends, geographical patterns and research needs', Preslia 80, 101-149. [ Links ]
Lantschner, M.V., Villacide, J.M., Garnas, J.R., Croft, P., Carnegie, A.J., Liebhold, A.M. et al., 2014, 'Temperature explains variable spread rates of the invasive woodwasp Sirex noctilio in the Southern Hemisphere', Biological Invasions 16, 329-339. https://doi.org/10.1007/s10530-013-0521-0 [ Links ]
Latombe, G., Pyšek, P., Jeschke, J.M., Blackburn, T.M., Bacher, S., Capinha, C. et al., in press, 'A vision for global monitoring of biological invasions', Biological Conservation. https://doi.org/10.1016/j.biocon.2016.06.013 [ Links ]
Lever, C., 1987, Naturalized birds of the world, Longman Scientific and Technical, Essex. [ Links ]
Lever, C., 2005, Naturalised birds of the world, T & A D Poyser, London. [ Links ]
Lockwood, J.L., Cassey, P. & Blackburn, T.M., 2005, 'The role of propagule pressure in explaining species invasions', Trends in Ecology and Evolution 20(5), 223-228. https://doi.org/10.1016/j.tree.2005.02.004 [ Links ]
Lombaert, E., Guillemaud, T., Cornuet, J.-M., Malausa, T., Facon, B. & Estoup, A., 2010, 'Bridgehead effect in the worldwide invasion of the biocontrol harlequin ladybird', PLoS One 5(3), e9743. https://doi.org/10.1371/journal.pone.0009743 [ Links ]
Long, J.L., 1981, Introduced birds of the world, David & Charles, London. [ Links ]
Lucy, F.E., Roy, H.E., Simpson, A., Carlton, J.T., Hanson, J.M., Magellan, K. et al., 2016, 'INVASIVESNET towards an international association for open knowledge on invasive alien species', Management of Biological Invasions 7(2), 131-139. https://doi.org/10.3391/mbi.2016.7.2.01 [ Links ]
Matthews, S. & Brand, K., 2004, Africa invaded: The growing danger of invasive alien species, The Global Invasive Species Programme, Cape Town. [ Links ]
McGeoch, M.A., Butchart, S.H.M., Spear, D., Marais, E., Kleynhans, E.J., Symes, A. et al., 2010, 'Global indicators of biological invasion: Species numbers, biodiversity impact and policy responses', Diversity and Distributions 16, 95-108. https://doi.org/10.1111/j.1472-4642.2009.00633.x. [ Links ]
Measey, J., Davies, S., Vimercati, G., Rebelo, A., Schmidt, W. & Turner, A., 2017, 'Invasive amphibians in southern Africa: A review of invasion pathways', Bothalia 47(2), a2117. https://doi.org/10.4102/abc.v47i2.2117 [ Links ]
Muatinte, B.L., van den Berg, J. & Santos, L.A., 2014, 'Prostephanus truncatus in Africa: A review of biological trends and perspectives on future pest management strategies', African Crop Science Journal 22(3), 237-256. [ Links ]
Peacock, D.S., van Rensburg, B.J. & Robertson, M.P., 2007, 'The distribution and spread of the invasive alien common myna, Acridotheres tritis L (Aves: Sturnidae), in southern Africa', South African Journal of Science 103, 465-473. [ Links ]
Picker, M. & Griffiths, C.L., 2011, Alien and invasive animals: A South African perspective, Struik Nature, Cape Town. [ Links ]
Poutsma, J., Loomans, A.J.M., Aukema, B. & Heijerman, T., 2008, 'Predicting the potential geographical distribution of the harlequin ladybird, Harmonia axyridis, using the CLIMEX model', BioControl 53, 103-125. https://doi.org/10.1007/s10526-007-9140-y [ Links ]
Roques, A., Auger-Rozenberg, M.-A., Blackburn, T.M., Garnas, J., Pyšek, P., Rabitsch, W. et al., 2016, 'Temporal and interspecific variation in rates of spread for insect species invading Europe during the last 200 years', Biological Invasions 18(4), 907-920. https://doi.org/10.1007/s10530-016-1080-y [ Links ]
Rouget, M., Robertson, M.P., Wilson, J.R.U., Hui, C., Essl, F., Renteria, J.L. et al., 2016, 'Invasion debt - Quantifying future biological invasions', Diversity and Distributions 22, 445-456. https://doi.org/10.1111/ddi.12408 [ Links ]
Roy, H.E., Brown, P.M.J., Adriaens, T., Berkvens, N., Borges, I., Clusella-Trullas, S. et al., 2016, 'The harlequin ladybird, Harmonia axyridis: Global perspectives on invasion history and ecology', Biological Invasions 18(4), 997-1044. https://doi.org/10.1007/s10530-016-1077-6 [ Links ]
Secretariat of the Convention on Biological Diversity, 2010, 'Guide to the global taxonomy initiative', CBD Technical Series, viewed 29 June 2016, from https://www.cbd.int/doc/publications/cbd-ts-30.pdf [ Links ]
Seebens, H., Essl, F., Dawson, W., Fuentes, N., Moser, D., Pergl, J. et al., 2015, 'Global trade will accelerate plant invasions in emerging economies under climate change', Global Change Biology 21(11), 4128-4140. https://doi.org/10.1111/gcb.13021. [ Links ]
Skelton, P., 1993, A complete guide to the freshwater fishes of Southern Africa, Southern Book Publishers, Harare. [ Links ]
Slippers, B., Wingfield, M.J., Coutinho, T.A. & Wingfield, B.D., 2001, 'Population structure and possible origin of Amylostereum areolatum in South Africa', Plant Pathology 50, 206-210. https://doi.org/10.1046/j.1365-3059.2001.00552.x [ Links ]
Slippers, B., Wingfield, B.D., Coutinho, T.A. & Wingfield, M.J., 2002, 'DNA sequence and RFLP data reflect geographical spread and relationships of Amylostereum areolatum and its insect vectors', Molecular Ecology 11, 1845-1854. https://doi.org/10.1046/j.1365-294X.2002.01572.x [ Links ]
Small, E., 2011, Alfalfa and relatives: Evolution and classification of Medicago, NRC Research Press, Ottawa. [ Links ]
Stals, R. & Prinsloo, G., 2007, 'Discovery of an alien invasive, predatory insect in South Africa: The multicoloured Asian ladybird beetle, Harmonia axyridis (Pallas) (Coleoptera: Coccinellidae)', South African Journal of Science 103, 123-126. [ Links ]
Stals, R., 2010, 'The establishment and rapid spread of an alien invasive lady beetle: Harmonia axyridis (Coleoptera: Coccinellidae) in southern Africa, 2001-2009', IOBC/wprs Bulletin 58, 125-132. [ Links ]
Taylor, J.S., 1962, 'Sirex noctilio F., a recent introduction in South Africa', Entomologist's Record 74, 273-274. [ Links ]
Tribe, G.D., 1995, 'The woodwasp Sirex noctilio Fabricius (Hymenoptera: Siricidae), a pest of Pinus species, now established in South Africa', African Entomology 3(2), 215-217. [ Links ]
Tribe, G.D. & Cilliè, J.J., 2004, 'The spread of Sirex noctilio Fabricius (Hymenoptera: Siricidae) in South African pine plantations and the introduction and establishment of its biological control agents', African Entomology 12(1), 9-17. [ Links ]
UNEP, 2011, Report of the tenth meeting of the conference of parties to the Convention on Biological Diversity, Nagoya, Japan, 18-29 October 2010. [ Links ]
van Kleunen, M., Dawson, W., Essl, F., Pergl, J., Winter, M., Weber, E. et al., 2015, 'Global exchange and accumulation of non-native plants', Nature 525, 100-103. https://doi.org/10.1038/nature14910 [ Links ]
van Rensburg, B.J., Weyl, O.L.F., Davies, S.J., van Wilgen, N.J., Spear, D., Chimimba, C.T. et al., 2011, 'Invasive vertebrates of South Africa', in D. Pimentel (ed.), Biological invasions: Economic and environmental costs of alien plant, animal, and microbe species, pp. 325-378, CRC Press, Boca Raton, FL. [ Links ]
Visser, V., Wilson, J.R.U., Canavan, S., Fish, L., Le Maitre, D.C., Nänni, I. et al., 2017, 'Grasses as invasive plants in South Africa revisited: Patterns, pathways and management', Bothalia 47(2), a2169. https://doi.org/10.4102/abc.v47i2.2169 [ Links ]
Wells, M.J., Balsinhas, A.A., Joffe, H., Engelbrecht, V.M., Harding, G. & Stirton, C.H., 1986, 'A catalogue of problem plants in Southern Africa', Memoirs of the Botanical Survey of South Africa 53, 1-658. [ Links ]
Willis, C.K. & Huntley, B.J., 2001, 'Developing capacity within southern Africa's herbaria and botanical gardens', Systematics and Geography of Plants 71(2), 247-258. https://doi.org/10.2307/3668671 [ Links ]
Wilson, J.R.U., Dormontt, E.E., Prentis, P.J., Lowe, A.J. & Richardson, D.M., 2009, 'Something in the way you move: Dispersal pathways affect invasion success', Trends in Ecology and Evolution 24(3), 136-144. https://doi.org/10.1016/j.tree.2008.10.007 [ Links ]
Wilson, J.R.U., Gaertner, M., Richardson, D.M. & van Wilgen, B.W., 2017, 'Contributions to the National Status Report on Biological Invasions in South Africa', Bothalia 47(2), a2207. https://doi.org/10.4102/abc.v47i2.2207 [ Links ]
Wingfield, M.J., Slippers, B., Hurley, B.P., Coutinho, T.A., Wingfield, B.D. & Roux, J., 2008, 'Eucalypt pests and diseases: Growing threats to plantation productivity', Southern Forests 70(2), 139-144. https://doi.org/10.2989/SOUTH.FOR.2008.70.2.9.537 [ Links ]
Zuur, A.F., Ieno, E.N., Walker, N.J., Saveliev, A.A. & Smith, G.M., 2009, Mixed effects models and extensions in ecology with R, Springer, New York. [ Links ]
 Correspondence:
Correspondence:
Katelyn Faulkner
katelynfaulkner@gmail.com
Received: 30 Aug. 2016
Accepted: 10 Dec. 2016
Published: 31 Mar. 2017
Note: This paper was initially delivered at the 43rd Annual Research Symposium on the Management of Biological Invasions in South Africa, Goudini Spa, Western Cape, South Africa on 18-20 May 2016.
Appendix 1







{kind=link}
{kind=link}
{kind=link}
{kind=link}