










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.47 n.1 Pretoria 2017
http://dx.doi.org/10.4102/abc.v47i1.2260
SHORT COMMUNICATION
Status of the Genera Colpoon, Osyris and Rhoiacarpos in South Africa
Daniel L. Nickrent
Department of Plant Biology, Southern Illinois University, United States
ABSTRACT
BACKGROUND: The taxonomic and phylogenetic status of Colpoon, Osyris and Rhoiacarpos (Santalaceae, Osyrideae) is reviewed.
OBJECTIVES: To resolve confusion regarding whether Colpoon is deserving of generic status separate from Osyris.
METHODS: Existing morphological information was examined for the three genera as well as previously published molecular phylogenies.
RESULTS: From both morphological and phylogenetic perspectives, Colpoon is distinct from Osyris. The status of Rhoiacarpos was not contentious and this genus is also easily differentiated from the other two genera in Osyrideae.
CONCLUSIONS: Colpoon and Osyris are not congeneric; therefore, floras, databases and herbarium collections should recognise these as distinct taxa.
Introduction
Over the past several decades, some confusion has surrounded the taxonomic circumscription of three genera of South African Santalaceae, tribe Osyrideae: Osyris L. (1753), Colpoon P.J.Bergius (1767) and Rhoiacarpos A.DC (1857). The taxonomic history and generic boundaries (from a morphological perspective) of these three taxa were discussed by Stauffer (1961). For several decades following Stauffer's work, these three genera were generally treated as distinct, as shown in 'Plants of Southern Africa: Names and Distribution' by Arnold and De Wet (1993). In 1994, Hilliard published a one-page note on Colpoon where she followed the treatment in Flora Capensis (Hill 1915) and reduced Colpoon compressum P.J.Bergius to a synonym of Osyris abyssinica Hochst. ex A.Rich. (= O. lanceolata Hochst. & Steud.). As stated by Hilliard (1994), 'There is, however, no essential differences in floral detail or in the structure of the inflorescence; those given by Stauffer (1961) are illusory'. But are the differences between the three genera given in Stauffer (1961, Table 1) real or imagined? It should be noted that Stauffer examined specimens, including the types, from Zurich, Geneva, Paris, London and Kew. His work was the most comprehensive up to that time and he concluded that the three species 'can be clearly distinguished morphologically and among themselves show no transitions'. Most web sites, recent floras and field guides that appeared after Hilliard (1994) accepted two genera for South Africa, Osyris and Rhoiacarpos, with Colpoon listed as a synonym of the former. Examples include Goldblatt and Manning (2000), Bean and Johns (2005), Germishuizen et al. (2006), Mucina and Rutherford (2006) and Manning (2007). This trend was reversed by Manning and Goldblatt (2012) who took into account molecular phylogenetic data that showed the three genera were distinct. Despite this publication, the concept of two Osyrideae genera for South Africa persists in herbarium collections and both popular and scientific works. Therefore, the purpose of this paper is to summarise the existing morphological and molecular information that supports recognising three distinct genera.
Molecular phylogenetic data
The first comprehensive molecular phylogenetic study of Santalaceae was by Der and Nickrent (2008) that used nuclear small-subunit ribosomal DNA and chloroplast rbcL and matK. A portion of the tree there referred to as the 'Santalum clade' (now Santalaceae s. str.) is shown in Figure 1a. The clade composed of Osyris, Nestronia, Rhoiacarpos and Colpoon (tribe Osyrideae) is strongly supported as monophyletic. Moreover, Osyris quadripartita (Europe) was not sister to Colpoon, with Rhoiacarpos capensis occupying that position. It is relevant that Stauffer (1961) also saw this association based on morphology:
Therefore, an association of Colpoon and Osyris can not be accepted. If anything, one finds understanding of the union of Colpoon and Rhoiacarpos according to the actions of Baillon and Bentham. (p. 392)
One could argue that because no South African accession of Osyris was included, this result should not be used to address generic delimitation in this geographic region. A seven-gene study that sampled nuclear, chloroplast and mitochondrial genes from 197 members of Santalales was published (Su et al. 2015) and a portion of the resulting tree is shown in Figure 1b. In that study, Osyris lanceolata from South Africa was included and it is sister to the European Osyris quadripartita. As before, Osyris is sister to a clade containing Nestronia, Rhoiacarpos and Colpoon with the latter two sisters.
Morphological data
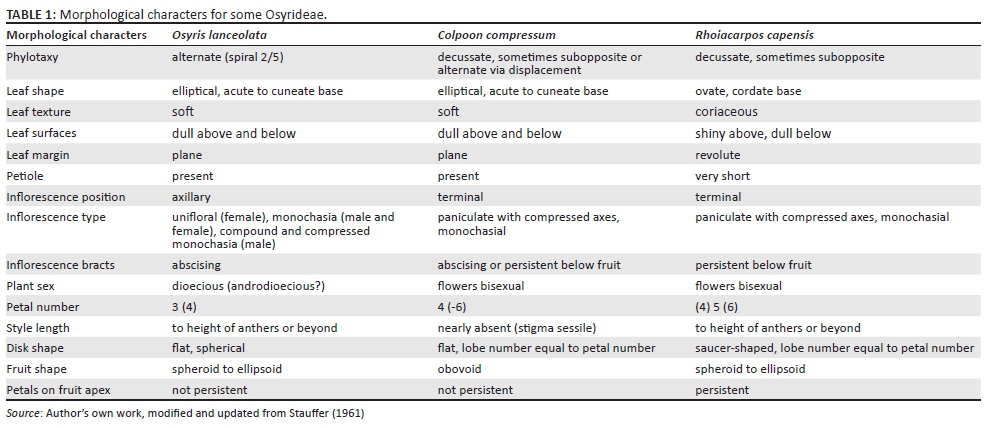
A comparison of the vegetative and reproductive morphologies of Osyris, Colpoon and Rhoiacarpos shows that these genera share a number of features and yet differ in others (Table 1 and Figures 2-4).
Leaves
All three genera have simple, entire leaves with mucronate apices. All show winged or ridged stems with the ridges merging with leaf petiole bases. Leaf phylotaxy was used by Hill (1915) to justify merging Colpoon with Osyris because both alternate and opposite leaves were seen in the former. Variation in phylotaxy was noted by Stauffer (1961) where C. compressum was scored as decussate, rarely with the two leaves somewhat shifted. Bean (1990) scored C. compressum as 'leaves usually opposite to subopposite, less often alternate' and the newly described species Colpoon speciosum as 'leaves as often alternate as subopposite, less often opposite'. Although Rhoiacarpos leaves were scored as opposite by Hill (1915), it too can show the subopposite condition (Figure 4b). Thus, phylotaxy appears to be a rather plastic morphological feature in Osyrideae. Shifting (displacement) of leaf position through development can be a form of metatopy (Weberling 1989) possibly influenced by genetic or environmental factors.
Rhoiacarpos is easily distinguished from the other two genera by leaf features. Its leaves are nearly sessile, bifacial (shiny above and dull below), with cordate bases and somewhat revolute margins. The leaves of Osyris and Colpoon differ in these characters, but are quite similar to each other overall. Hilliard (1994) states for both Osyris and Colpoon 'both leaf surfaces closely and minutely white-dotted', but this feature has not been observed by this author in the field, herbarium or in photographs.
Inflorescences
The inflorescences of Osyris are mainly axillary whereas those of Colpoon and Rhoiacarpos are terminal. It is interesting that Hill (1915) describes the inflorescence of Rhoiacarpos as a panicle composed of 3-flowered axillary cymules. In Figure 4 of Stauffer (1961), both Rhoiacarpos and Colpoon are diagrammed with 3-flowered units. In other Santalales, this 3-flowered cymule unit is equivalent to a dichasium. But the situation in Osyrideae appears more complex, mainly owing to compression and reduction (loss of flowers). Hilliard (1994) indicates flowers in Osyris lanceolata 'may occur in 2-3-flowered dichasia'. Technically, a dichasium is 3-flowered whereas a monochasium is 2-flowered. Upon first examination, the inflorescence units may appear to be dichasial such as in Osyris (Figure 2c), Colpoon (Figure 3b), and Rhoiacarpos (Figure 4c), but in these cases the pair of flowers (or buds) subtending the older, terminal flower are unequal in age. Moreover, the older of the two subtending buds is often present as a lateral innovation. For these reasons, it seems best to describe these units as monochasia or compound monochasia. Simple (2-flowered) monochasia are frequently seen in the female inflorescences of Osyris (Figure 2d).
The terminal peduncles in the panicles of Colpoon are often swollen (Figure 3a and d). In his diagram, Stauffer (1961) coloured black those inflorescence axes that become compressed. Thus, the cluster of flowers seen arising from these swollen apices in Colpoon is interpreted as being a series of reduced monochasia (or dichasia according to Stauffer 1961). The phenomenon of syndesmy (incorporation of partial inflorescence units and their axes into the main axis) is well documented in cymoid types resulting in a coenosome (Weberling 1989). Looking at the lower inflorescence branch in Figure 3a, the six flowers and buds are of different ages: two past anthesis, one at anthesis, one older bud and two younger buds. This situation could be interpreted as two dichasia, but because the ages of the flowers are wrong, this situation might best be interpreted as three monochasia.
The components of the inflorescence axes (peduncles) are often subtended by small bracts. Stauffer (1961) indicated that these abscise in Osyris, abscise or are persistent in Colpoon and are persistent in Rhoiacarpos. The study by Bean (1990) confirms that the persistence of these bracts in C. compressum is polymorphic (compare Figure 3a and d) and further shows that in C. speciosum the bracts are persistent and enlarge upon fruiting.
Flowers
The flowers in Colpoon and Rhoiacarpos are bisexual, whereas in Osyris lanceolata flowers appear to be unisexual. Hilliard (1994) stated, 'the flowers of Osyris lanceolata are hermaphrodite and male, the plants being androdioecious', but no evidence was provided that supported the presence of bisexual versus female flowers. Female flowers possess stamens (Figure 2d), but if the anthers can be shown to not produce pollen, then these are staminodes and the flower is functionally female.
Flower merosity is notoriously variable in Santalaceae, often showing three or four different forms within the same inflorescence. But for differentiating the three genera considered here, this character has some value when the most common number is considered. Thus, Osyris generally has 3-merous flowers, occasionally having 4-merous flowers. Colpoon typically has 4-merous flowers, but 5- and 6-merous flowers can also be seen. Most Rhoiacarpos flowers are 5-merous, but 4- and 6-merous flowers are also known. The number of lobes of the glandular disk in these genera follows the number of petals.
Fruits
In all three genera, many fewer fruits develop to maturity compared with the number of flowers originally present on the inflorescence axis. In Colpoon, evidence of flower abortion exists (Figure 3d) such that a single fruit is present per axis with the scars of abortive flowers present at the swollen peduncle apex (Figure 3f). The fruits of Rhoiacarpos differ from the other two genera in having persistent petals at the apex (Figure 4f and g). A cluster of fruits is shown in Figure 4f; however, each fruit appears to be present on a separate inflorescence branch with evidence of numerous abortive flowers and their associated bracts below (Figure 4g).
In Osyris (Figure 2f) and C. compressum (Figure 3f), the petals abscise and are not persistent on the mature fruit. Bean (1990) indicates that in C. speciosum the petals are persistent; thus, this feature is polymorphic within the genus. Although it is not clear whether the character of fruit shape will remain valid following further observations, it appears that the fruit in Colpoon is more obovoid than in the other two genera that have spheroid to ellipsoid fruits.
Discussion
The decision by Hilliard (1994) to lump Colpoon into Osyris can now be addressed given the molecular phylogenetic and comparative morphology results. From the topologies of the phylogenetic trees (Figure 1a and b), one could include Colpoon in Osyris. If this was done, and monophyly was maintained, then the North American Nestronia and South African Rhoiacarpos would also have to be included in Osyris. This concept has never been proposed and in fact Rhoiacarpos has been considered a separate genus in all consulted works on South African flora. Molecular dating conducted by Vidal-Russell and Nickrent (2008) showed that Santalum and Osyris diverged in the Paleocene, at least 60 million years ago. Although the other genera (Nestronia, Rhoiacarpos and Colpoon) have not been included in ultrametric trees, it is likely that they diverged from each other somewhat more recently, for example, in the Eocene, ca. 50 million years ago.
Despite the statement by Hill (1915) that 'There is also no floral difference between Colpoon Berg. and Osyris Linn'. and the similar sentiment expressed by Hilliard (1994), the evidence presented above shows that there are clear differences in vegetative, floral and fruit features among all three genera. The taxonomic conclusions reached by Stauffer (1961) are fully supported by molecular phylogenetic data; thus, Colpoon should be considered a genus, distinct from Osyris, and is composed of two species, C. compressum and C. speciosum.
Acknowledgements
I thank Marinda Koekemoer for allowing me to use several photos of Osyris lanceolata and Colpoon compressum. Photos of Rhoiacarpos capensis, posted online at www.floraoftheworld.org, were taken by Christopher Davidson. One of the reviewers is acknowledged for pointing out overlooked but important literature.
Competing interests
The author declares that he has no financial or personal relationships that may have inappropriately influenced him in writing this article.
References
Arnold, T.H. & De Wet, B.C., 1993, Plants of Southern Africa: Names and distribution, National Botanical Institute, Pretoria. [ Links ]
Bean, A. & Johns, A., 2005, Stellenbosch to Hermanus, South African Wild Flower Guide 5, Botanical Society of South Africa, Cape Town. [ Links ]
Bean, P.A., 1990, 'The identity of Osyris abyssinica var. speciosa (Santalaceae)', South African Journal of Botany 56, 665-669. https://doi.org/10.1016/S0254-6299(16)31004-3 [ Links ]
Der, J.P. & Nickrent, D.L., 2008, 'A molecular phylogeny of Santalaceae (Santalales)', Systematic Botany 33, 107-116. https://doi.org/10.1600/036364408783887438 [ Links ]
Germishuizen, G., Meyer, N.L., Steenkamp, Y. & Keith, M., 2006, A checklist of South African plants, SABONET Report no. 41, Southern African Botanical Diversity Network, Pretoria. [ Links ]
Goldblatt, P. & Manning, J.C., 2000, 'Cape plants: A conspectus of the Cape flora of South Africa', Strelitzia 9, 743. [ Links ]
Hill, A.W., 1915, 'Santalaceae', in W.T. Thiselton-Dyer (ed.), Flora Capensis, pp. 135-212, Vol. 5, L. Reeve & Co., Ltd., London. [ Links ]
Hilliard, O.M., 1994, 'A note on Colpoon (Santalaceae)', Edinburgh Journal of Botany 51, 391-392. https://doi.org/10.1017/S0960428600001840 [ Links ]
Manning, J., 2007, Field guide to fynbos, Struik (Division of New Holland Publishing), Cape Town. [ Links ]
Manning, J.C. & Goldblatt, P., 2012, 'Plants of the Greater Cape Floristic Region 1: The Core Cape flora', Strelitzia 29, 853. [ Links ]
Mucina, L. & Rutherford, M.C. (eds.), 2006, The vegetation of South Africa, Lesotho and Swaziland, South African National Biodiversity Institute, Pretoria. [ Links ]
Stauffer, H.U., 1961, 'Santalales-Studien V. Afrikanische Santalaceae I. Osyris, Colpoon und Rhoiacarpos', Vierteljahrsschrift der Naturforschenden Gesellschaft in Zurich 106, 387-400. (A German to English translation of this work is available on Parasitic Plant Connection at http://parasiticplants.siu.edu/Translations.html). [ Links ]
Su, H.-J., Hu, J.-M., Anderson, F.E. & Nickrent, D.L., 2015, 'Phylogenetic relationships of Santalales with insights into the origins of holoparasitic Balanophoraceae', Taxon 64, 491-506. https://doi.org/10.12705/643.2 [ Links ]
Vidal-Russell, R. & Nickrent, D.L., 2008, 'The first mistletoes: Origins of aerial parasitism in Santalales', Molecular Phylogenetics and Evolution 47, 523-527. https://doi.org/10.1016/j.ympev.2008.01.016 [ Links ]
Weberling, F., 1989, Morphology of flowers and inflorescences, English transl. R.J. Pankhurst, Cambridge University Press, Cambridge. [ Links ]
 Correspondence:
Correspondence:
Daniel Nickrent
nickrent@plant.siu.edu
Received: 16 May 2017
Accepted: 10 Oct. 2017
Published: 13 Nov. 2017


{kind=link}
{kind=link}