










Services on Demand
Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
Indicators

Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkBothalia - African Biodiversity & Conservation
On-line version ISSN 2311-9284
Print version ISSN 0006-8241
Bothalia (Online) vol.45 n.1 Pretoria 2015
http://dx.doi.org/10.4102/abc.v45i1.178
ORIGINAL RESEARCH
Impacts of fire and grazing management on South Africa's moist highland grasslands: A case study of the Steenkampsberg Plateau, Mpumalanga, South Africa
Ian T. LittleI, II; Philip A.R. HockeyI, †; Raymond JansenIII
IPercy FitzPatrick Institute of African Ornithology, University of Cape Town, South Africa
IIEndangered Wildlife Trust, Johannesburg, South Africa
IIIDepartment of Environmental, Water and Earth Sciences, Tshwane University of Technology, South Africa
ABSTRACT
BACKGROUND: Grasslands are heavily utilised for livestock agriculture and the resultant degradation through mismanagement contributes to an estimated 60% of this biome being permanently transformed. This study focused on the impact of fire and grazing in moist highland grasslands
OBJECTIVES: To determine the contribution of burning frequency and grazing intensity combined (for domestic livestock and indigenous ungulates) on vegetation structure heterogeneity and species diversity
METHODS: Eight study sites under different management regimes were sampled over two summers. Vegetation structure characteristics and diversity data were collected monthly within multiple replicates in each study site. A disc pasture meter was used to assess standing biomass. Differences in vegetation structure characteristics, plant community composition and plant species assemblage structure across sites were statistically analysed using analyses of variance, indicator species analyses, multidimensional scaling ordinations and two-way cluster analyses
RESULTS: The combination of heavy grazing and annual burning leads to a distinct plant community dominated by disturbance specialist species. Selective grazing by indigenous herbivores promotes a community of unpalatable species. This study illustrates that fenced indigenous herbivores, even at moderate stocking densities, have a greater detrimental impact on plant diversity and structure than do domestic livestock
CONCLUSION: Intensive grazing and burning have a detrimental impact on plant species diversity and structure. This also affects resultant palatability for grazing livestock and fenced game. To promote both grazing quality and ecological integrity we recommend a minimum sustainable 'fodder capacity' or standing phytomass of 5000 kg per large-animal unit per hectare for domestic livestock in moist highland grasslands
Introduction
Grasslands are ancient, complex and slowly evolving systems of diverse plant communities that generally lack the ability to recover after severe disturbance (Jacobs, Kingston & Jacobs 1999; Little et al. 2005). South African grasslands are being increasingly degraded through the cumulative influence of overgrazing (Neke & Du Plessis2004; O'Connor et al. 2011), extensive, frequent burning (Uys, Bond & Everson 2004), plantation forestry (Allan et al. 1997; Lipsey & Hockey 2010) and invasion by alien plant species (Le Maitre et al. 1996). In South Africa, it is estimated that 60% of the grassland biome has been permanently transformed, whilst as little as 15% remains as natural grassland. Of the grassland biome, 25% is degraded to some degree and only about 2% is formally conserved (Carbutt et al. 2011; Macdonald 1989). Of particular concern is that the majority of the remaining natural grassland is highly fragmented and most is poorly managed (Mucina & Rutherford 2006:362; South African National Biodiversity Institute 2014). Grasslands have traditionally been used for livestock farming, which has allowed some of the natural ecological processes to continue fairly undisturbed. However, livestock grazing does not simulate the natural herbivore-grassland interaction that would have occurred in the area historically, at least from the late Miocene and early Pliocene (Jacobs et al. 1999; McNaughton 1986), and impacts of intensive grazing on species diversity in this system are poorly understood (Uys et al. 2004).
The use of grazing, and more commonly fire, for managing grassland is primarily to provide a green flush of nutrient-rich grass, remove old moribund grass material and control tick abundance (Bond 1997; Van Wilgen & Scholes 1997). However, grazing by livestock and the associated trampling and nutrient enrichment have a significant impact on grassland plant diversity (O'Connor et al. 2010; O'Connor et al. 2011; Veen et al. 2008), as does the use of fire (Veen et al. 2008). Furthermore, selective grazing has been shown to alter the structure and species composition of grasslands (Milchunas, Sala & Lauenroth 1988; Owen-Smith 1999), often favouring unpalatable species, which, in turn, causes a change in community species composition (Milchunas et al. 1988; Sternberg et al. 2000). Fire also modifies the grazing pattern and behaviour of herbivores, because it reduces above-ground biomass (Van Wilgen & Scholes 1997).
Although South African grassland systems are naturally maintained by winter and spring fires (Mucina & Rutherford 2006:356), there is concern over the possible detrimental effect of unnaturally frequent fires on plant diversity (Collins & Calabrese 2012; Swengel 2001). It has been suggested that the natural fire frequency in moist highland grasslands would have been only every 4 years or more (Manry & Knight 1986). It has further been suggested that controlled burning should be based on the rate of litter accumulation and that grazing should not start until sward height reaches 250 mm (Mentis 1981). This is manageable with domestic livestock, but indigenous game species tend to graze burnt areas selectively and therefore their stocking rates in fenced areas need to be sufficiently low so that the total number of animals congregating on burnt areas alone is not detrimental (Van Rooyen, Bredenkamp & Theron 1986). Annual burning of grasslands after the first rains has become common practice amongst farmers (Everson 1999), irrespective of litter accumulation (Everson 1999; Van Wilgen & Scholes 1997). However, without frequent burning, some species (e.g. Themeda triandra Forssk 1775 and Heteropogon contortus Roem & Schult 1954) become moribund and decrease in abundance, whereas other species are more tolerant of self-shading and increase (Owen-Smith & Danckwerts 1997).
Tainton (1999) has suggested that the carrying capacity for high-altitude climax grassland lies somewhere between 3 ha and 5 ha per large-animal unit (ha LAU−1). One LAU is defined as being equivalent to one cow or five sheep and represents the metabolic equivalent of a 454-kg cow (Owen-Smith & Danckwerts 1997; Tainton 1999). However, many farmers exceed this stocking rate and thereby exceed the ecological carrying capacity of the grasslands. Together with annual burning the system's ecological integrity and future livestock productivity are likely to be adversely affected (Fuhlendorf et al. 2012; Limb et al. 2011; Little, Hockey & Jansen 2013).
Historically, indigenous game species in moist highland grasslands moved seasonally along rainfall gradients, which resulted in a temporal and spatial easing of the grazing pressure (Rowe-Rowe & Scotcher 1986). Indeed, there probably was no grazing by indigenous game species in high-altitude grasslands during the dry winter months (Mucina & Rutherford 2006:357). The effects of indigenous game species on plant diversity under current management practices have yet to be investigated.
Previous studies on the effect of fire (Uys et al. 2004) and grazing (Owen-Smith & Danckwerts 1997) on the grassland biome in South Africa have focused on their independent impacts, with limited attention given to their combined effect. O'Connor et al. (2010) highlighted the need to assess the combined effect on plant communities. Previous studies in moist grasslands have not included a detailed analysis of the influence of grazing regimes or burning frequency on vegetation structure, which may have a profound effect on grassland ecosystem functioning. Therefore, the objective of this study was to assess the interactive effect of burning frequency and stocking rates on plant community (vegetation) structure and diversity in moist highland grasslands. It is hypothesised that with increased stocking rates plant diversity will decrease, accompanied by a shift from predominantly decreaser to predominantly increaser grasses (Tainton 1999). Decreasers are species that are sensitive to degraded grassland conditions as a result of either too much or too little disturbance. Increaser I species increase in abundance as a result of too little disturbance (typically when the grassland becomes moribund), increaser II species increase in abundance in response to heavy disturbance (e.g. by fire, grazing or a combination), and increaser III species proliferate in response to selective grazing (i.e. are unpalatable species - Tainton [1999]). The study also compares the impacts of indigenous game (predominantly blesbok [Damaliscus dorcas]) to those of (more heavily stocked) domestic livestock on vegetation structure and diversity.
Research method and design
Study sites
The study area falls within the Moist Highland Grassland Bioregion and is specifically identified as Lydenburg Montane Grassland (Mucina & Rutherford 2006:405) (previously referred to as the North-Eastern Sandy Highveld grassland; Burgoyne, Bredenkamp & Van Rooyen [2000]). All the study sites were on the plateau around Dullstroom (centred at 25°25′S, 30°10′E) and were between 1900 m a.s.l. and 2200 m a.s.l. The soils are mostly derived from quartzite of the Steenkampsberg and Lakenvlei formations (Burgoyne et al. 2000). The Dullstroom Plateau Grassland (encompassing this entire study area) has been classified as endangered owing to very high irreplaceability of species (Department of Environmental Affairs 2011). The topography within the geographic range of this vegetation type includes high-altitude plateaus and undulating plains. The predominant vegetation is short grass in the high-lying areas, which becomes taller on the lower slopes. Although grass species diversity is relatively low, the diversity of non-graminoid elements of the vegetation ('forbs' from here on) is high (Mucina & Rutherford 2006:405). Orographic and convection-based precipitation (660 mm year−1 - 1180 mm year−1; Mucina and Rutherford [2006:405]) and heavy mists throughout most months of the year have promoted a unique flora, including a rich diversity of mesophytic plants such as the Orchidaceae. In fact, the high forb diversity (over 2260 taxa and 51 endemic species) has prompted this grassland type to be proposed as a 'centre of plant endemism' (Mucina & Rutherford 2006:360).
Eight study sites of similar soil type and underlying geology but under different management regimes (Table 1) were sampled over two summers. These sites (reported with their stocking rates) represent the most common types of management in the moist highland grasslands:
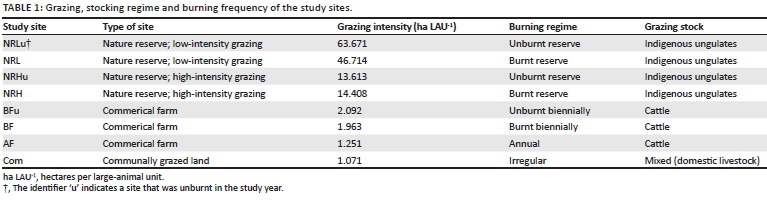
- an annually burnt farm (AF); 1.251 ha LAU−1
- communally grazed lands (Com); 1.071 ha∙LAU−1
- a biennially burnt farm that was either burnt (BF; 1.963 ha LAU−1) or left unburnt (BFu; 2.092 ha LAU−1) in the study year
- a nature reserve site, with a high density of indigenous grazing ungulates, which was either burnt (NRH; 14.408 ha LAU−1) or left unburnt (NRHu; 13.613 ha LAU−1) in the study year
- a nature reserve site, with a low density of indigenous grazing ungulates, whixh was either burnt (NRL; 46.715 ha LAU−1) or left unburnt (NRLu; 63.671 ha LAU−1) in the study year.
Numbers of grazing animals were constant throughout the season at all sites. The biennially burnt farm and both the heavily grazed and lightly grazed reserve sites had different stocking rates in the burnt and unburnt years; this was unavoidable and is a result of the flux based on mortality and fecundity. However, these changes in stocking rates between years were minimal and did not affect the categorisation of sites, and the burnt and unburnt sites can be considered as separate treatments. All nature reserve sites were within Verloren Valei Nature Reserve, a site that acted as both a control and a management experiment site in the heavily grazed area (NRH and NRHu). The reserve site was selected based on its good current condition and former management for conservation: the reserve was proclaimed in 1983 and has been block-burnt biennially since 1985 (Heyns 1985). All of the sites (except communally grazed lands where fire has not been used as a management tool for more than 50 years) are burnt in early spring after the first rains. Both the annually burnt and biennially burnt farms have been managed this way for at least three generations (> 100 years). The commercially farmed areas (AF and BF) were rotationally grazed by cattle on a four-day cycle by the same herd. In all cases the rotation was between two large camps. The stocking rate on communal lands (mixed grazing by cattle, sheep, horses, goats and pigs), where there is no stock rotation, is thus the number of animals in the area of both camps combined. Offspring of both livestock and indigenous grazers born in early summer were treated as 0.5 LAUs.
Within each study site, four 25-ha replicates were marked out with at least 500 m between replicates. Sampling plots and transects were randomly selected within each 500 m × 500 m replicate. Thus, plots and transects were nested within each of the four replicates, which, in turn, were nested within each of the eight study sites. As far as possible, these replicates were sufficiently far apart to avoid pseudo-replication but sufficiently close together to standardise for extrinsic factors such as geology, rainfall, aspect, slope and temperature within and between land-use treatments (Hurlbert 1984). The climatic aspects were comparable primarily owing to the limited altitudinal variation between sites (Burgoyne et al. 2000).
Fieldwork was conducted in the summer (October - March) for 2 years (2007-2009). Grazing data were collected by interviewing farmers or managers of chosen sites and were validated with animal counts. Relative grazing intensity was recorded as the area of grazing land available per LAU (ha LAU−1).
Vegetation structure
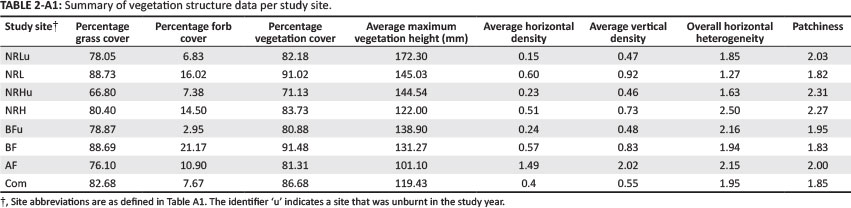
To assess the response of vegetation structure to management, it was important to understand which of the measured structural indices were most sensitive to the disturbance effects and which one could act as a surrogate for overall vegetation structure. When all 10 of the structural indices were included in the analysis (Table 2), any of the indices could have been used to differentiate between management practices but some were more effective than others. For this reason, a stepwise analysis was required to identify the most effective indices for sampling grassland structure.
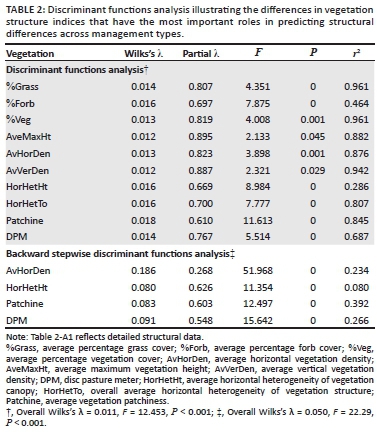
A backward stepwise discriminant functions analysis (Table 2) identified four of the original 10 structural indices as performing best at discriminating between management types. These were subsequently used in the multidimensional scaling plot analyses.
Vegetation structure was sampled monthly throughout the sampling season using two techniques. The first, modified from Wiens and Rotenberry (1981), involved establishing three 500-m transects within each of the four replicates per study site. Along each of these, at 50-m intervals, a 10-m long tape was laid out perpendicular to the transect line. Vegetation was sampled at 1-m intervals along the tape, yielding 100 point samples per transect (in total, 1200 samples per site per month). At each sampling point, a rod of 6-mm diameter was positioned vertically through the vegetation to the ground. This rod was marked at eight height intervals (0 mm - 50 mm, 50 mm - 100 mm, 100 mm - 200 mm, 200 mm - 300 mm, 300 mm - 400 mm, 400 mm - 500 mm, 500 mm - 600 mm and 600 mm - 1000 mm). At each sampling point the number of vegetation contacts with the rod per height interval was recorded, as well as whether the contact was with a grass or a forb. Open ground (in the case of no contact with vegetation) and overall maximum height of vegetation were also recorded. From these data we calculated three sets of vegetation structure indices (Table 2):
- Cover: Percentage grass cover (%grass), forb cover (%forb) and total plant cover in the form of grass and forbs combined (%veg), calculated as the percentage of point records with each of these parameters present.
- Structural measures: Average maximum height of vegetation (AveMaxHt) and horizontal density (AvHorDen), derived from the mean number of contacts with the rod in the 0 mm - 100 mm interval, and vertical density (AvVerDen), derived from the mean number of contacts over the entire length of the rod.
- Heterogeneity measures: Canopy heterogeneity (HorHetHt), given by the coefficient of variation of the maximum height contacts, overall heterogeneity (HorHetTo), given by the coefficient of variation of the mean total number of contacts over the entire rod, and a patchiness index (Patchine), in which the 10 samples per 50 m are grouped and then used to calculate the landscape patchiness according to the equation of Wiens and Rotenberry (1981):
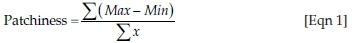
where Max represents the maximum number of contacts recorded in each sample group, Min represents the minimum number of contacts recorded in each sample group, and x represents the mean number of contacts recorded in each sample group.
The second technique quantifies vegetation density, which is a proxy for forage availability. Measurement of forage availability is essential for determining stocking rates for the management of herbivores (Ganguli et al. 2000; Smith 2006). A relative measure of forage availability was developed to assess how stocking density affected vegetation density through the season in a particular area. Vegetation density (kg ha−1) was combined with grazing intensity (ha LAU−1) to produce an index of available vegetation per LAU (kg LAU−1), which gives a clear indication of both the available above-ground biomass for grazing and the seasonal effect of grazing on vegetation biomass.
Vegetation density was sampled using a disc pasture meter (DPM), following Bransby and Tainton (1977), Danckwerts and Trollope (1980) and Trollope and Potgieter (1986). The DPM consists of a disc (1.5 kg, 457 mm in diameter) mounted onto a central rod, with a measuring scale set in 5-mm increments. The disc is dropped vertically from 0.6 m above the ground onto the grass sward and the settling height of the disc is recorded. Vegetation was sampled every 5 m along the three 500-m transects, yielding 100 DPM samples per transect and a total of 1200 samples per site per month.
The DPM is calibrated for a specific vegetation type to convert the reading into biomass estimates (kg ha−1). DPM calibrations are available for a number of vegetation types both within and outside of South Africa (Bransby & Tainton 1977; Danckwerts & Trollope 1980; Trollope & Potgieter 1986) for comparative purposes, but a callibration for moist highland grassland has not yet been done. To calibrate the DPM for moist highland grassland, monthly data were collected throughout the season from all the management types. The data were analysed using a regression analysis, with the standing crop of plant material harvested (removed and weighed) from below each disc measurement (converted to kg ha−1) as the dependent variable and the mean settling height of the disc as the independent variable. The regression analyses were repeated with the standing crop of grass subjected to logarithmic, square, square-root and reciprocal transformations to obtain the best linear fit between disc height and the standing crop of herbaceous plant material. The best fit was then compared with previous calibrations of the DPM.
Plant species diversity
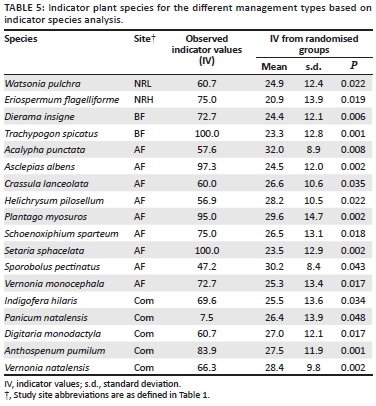
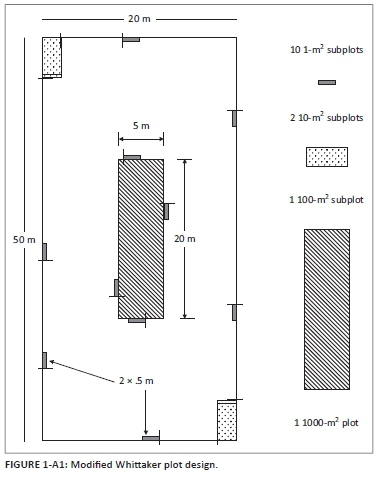
Plant species diversity in the five management types was assessed only in the years that sites were not burnt. Sampling was conducted in late January (when the majority of species were in flower), which allowed for species identification (Table 1-A1). Plants that could not be identified in the field were pressed and sent to the herbarium of the South African National Biodiversity Institute, Pretoria, for expert identification. Modified Whittaker nested vegetation sampling plots (O'Connor 2005; Figure 1-A1) were used to sample floral species diversity based on the methods of Stohlgren, Falkner and Schell (1995). Overall relative floral diversity assessments were based on one modified Whittaker plot per replicate (four per site). All species were recorded and abundance (percentage cover) of each species was estimated from each of the 10-m2 and 1-m2subplots. Estimated abundances per subplot were summed and the overall estimated abundance per species was calculated as a percentage. Plant species names were based on Germishuizen and Meyer (2003).
Statistical analyses
The software packages PC-ORD 5.10 (McCune & Mefford 2006) and STATISTICA 9.0 (StatSoft) were used for data analysis.
To assess the relative impacts of management practices on vegetation structure variables, analysis of variance (ANOVA) with post hoc Tukey tests were performed. A discriminant function analysis was subsequently undertaken, followed by a backward stepwise discriminant function analysis, which incorporated only the significant parameters, to extract the parameters that best revealed the effects of management on vegetation structure.
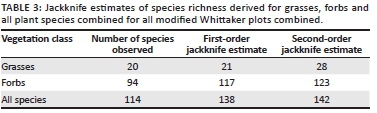
To assess sampling efficiency, species-area curves were fitted to the plant species richness data. First-and second-order jackknife estimates of species richness were derived separately for grasses, forbs and all species.
Plant community compositions were contrasted amongst management types using a one-way pairwise permutation-based non-parametric multivariate analysis of variance (PerManova; Anderson 2001; McCune & Mefford 2006). We used the Sørenson (Bray-Curtis) distance measure because it is appropriate for abundance data and gives robust outputs with zero-dominated datasets (McCune & Grace 2002). To assess the contribution of each species' abundance to the community and how much their response to disturbance has influenced each of these communities, we ran an indicator species analysis (McCune & Mefford 2006) using a Monte Carlo test of significance with 5000 permutations (Dufrêne & Legendre 1997); this method combines species' abundance and occurrence. A 'perfect indicator' should be present in all replicates within a site and not present in any other sites. To test whether grasses and forbs respond in a similar manner to site management, we performed a Mantel test (Douglas & Endler 1982), which tests the null hypothesis of no relationship between matrices and uses a t-distribution with infinite degrees of freedom. To test whether grass species experienced shifts from high to low palatability (based on Tainton [1999]) and from decreaser to increaser grasses with increasing disturbance, we ran Mann-Whitney U tests and ANOVA by ranks.
Multidimensional scaling (MDS) ordination was used in the analysis of vegetation structure between study sites as it is the statistical method of choice in community ecology when data are discontinuous or not normally distributed (McCune & Grace 2002). MDS was run (in PC-ORD) using a Euclidean distance measure with Bray-Curtis original endpoint selection for vegetation structure space and a Sørenson distance measure, which is recommended for community analyses, with Bray-Curtis original endpoint selection for plant species space. These ordinations fit matrix data into a two-dimensional space: in this case, the two data matrices analysed were vegetation structure diversity and species diversity (Bray & Curtis 1957; McCune & Grace 2002). DPM data were overlaid on the ordination and vegetation structure indices were included as a biplot.
Finally, to assess the shift in plant species assemblages as a result of land use, we ran a two-way cluster dendrogram (McCune & Mefford 2006) with a Sørenson distance measure and group-average linking method without relativisation.
Ethical considerations
This study was approved by the Scientific and Ethics Committee, University of Cape Town, and a plant collecting permit (MPB 1161) was obtained from the Mpumalanga Tourism and Parks Agency for the study period.
Results
Disc pasture meter calibration
The best calibration for moist highland grassland (locally known as 'sourveld') was linear (y = 358.7x - 746.4, with r = 0.95, r2 = 0.91; P < 0.0001) and had a higher r2value than calibrations derived from the other grassland systems, which rendered it a better fit. Nonetheless, existing calibrations (Bransby & Tainton 1977; Danckwerts & Trollope 1980; Trollope & Potgieter 1986) exhibit very similar trends to the moist highland grassland calibration.
Impact of management on vegetation structure
High densities of indigenous herbivores depleted grass and vegetation to the point that the NRH site (13.61 ha LAU−1) had both less grass cover (P < 0.01, df = 136) and less vegetation cover (P < 0.0005, df = 136) than any other site. This site also had the greatest vegetation patchiness (P < 0.01).
Biennial burning, coupled with heavy grazing by domestic stock, resulted in the BF site (2.09 ha LAU−1) having less forb cover than any other site (P < 0.05). Extremely heavy grazing, regardless of burning as occurs in the AF (1.25 ha LAU−1) and Com (1.07 ha LAU−1) sites, resulted in both higher horizontal vegetation density (P < 0.001 and P < 0.05, respectively) and lower vegetation biomass (P < 0.05 in both cases) than in any of the other sites.
The reserve site NRL (63.67 ha LAU−1), which is managed for conservation, had greater vegetation biomass (P < 0.05) and lower average horizontal vegetation density than any other site (P < 0.05).
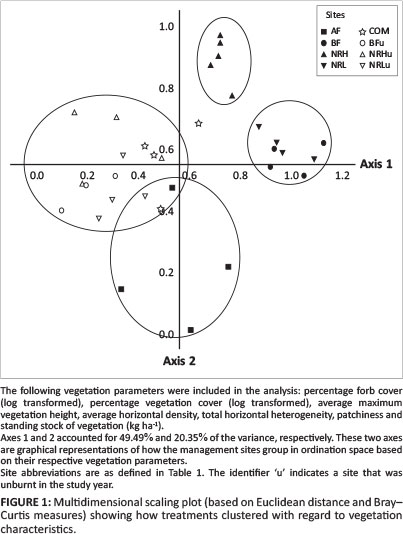
An ordination analysis was performed to gain a better understanding of how management type differentially affects vegetation structure (Figure 1). This allowed differentiation between management effects, including separating out the relative importance of fire frequency and stocking rate in influencing vegetation structure. Axes 1 and 2 accounted for 49.49% and 20.35% of the variance, respectively. (The two axes are graphical representations of how the management sites group in ordination space based on their respective vegetation parameters.)
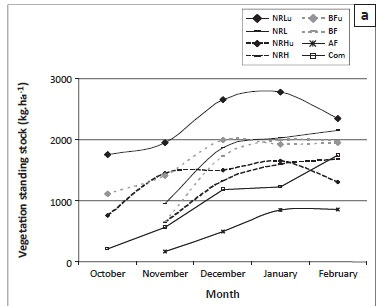
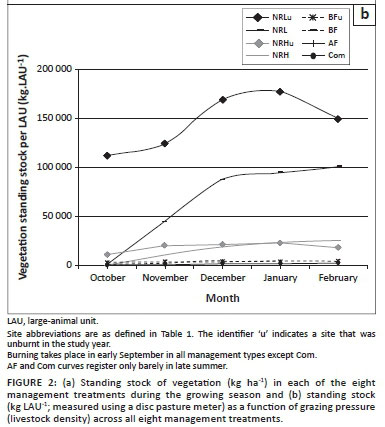
Grazing and burning directly influenced phytomass, which was highest in the site experiencing the lowest grazing pressure (NRL); this was true towards the end of the season even in the year that this site was burnt (Figure 2a).The BF site had the second-highest standing stocks and again, even in the year it was burnt, recovered to support a higher vegetation biomass than other commercially farmed sites. The AF and Com sites consistently had the lowest standing stocks of vegetation. However, intensive grazing by indigenous ungulates in the conservation area (NRH) resulted in lower phytomass by the end of the year than was present in the communally grazed lands. The results were similar for vegetation available per LAU (Figure 2b) for NRL, with low stocking rates.
Impact of management on plant species diversity
Species-area curves and jackknife estimates showed a small difference between observed and expected species richness (Table 3).
Between-site species diversity was assessed using PerManova (Table 4). A randomisation test of significance of pseudo F-values between all sites showed a significant overall difference between sites (F = 2.9022, P < 0.0005, df = 35).
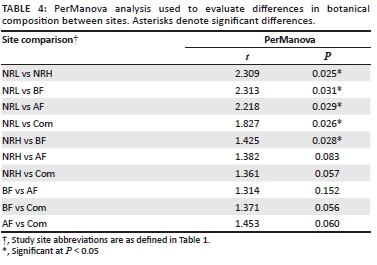
In support of the findings from the pairwise PerManova (Table 4), 37.7% of species occurred in only one management type (NRL). Only 3.51% and 7.89% of species were unique to NRH and BF, respectively. Two species found in the study area (Eucomis autumnalis Chit. 1951 and Hypoxis hemerocallidea Fisch 1842, both in AF and NRL) are currently threatened (Raimondo et al. 2009). One species, Plantago myosuros Lam. 1972 (in AF and Com), is an exotic weed from South America. A large number of increaser and pioneer species are present in the system (Table 5), as illustrated by the species that are confined to high disturbance areas, including the AF and Com sites. Two species were restricted to the conservation area and one of these was most abundant in the area that was heavily grazed by indigenous herbivores (NRH).
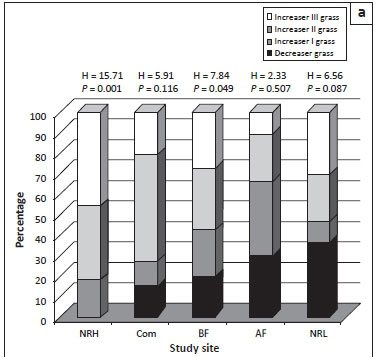
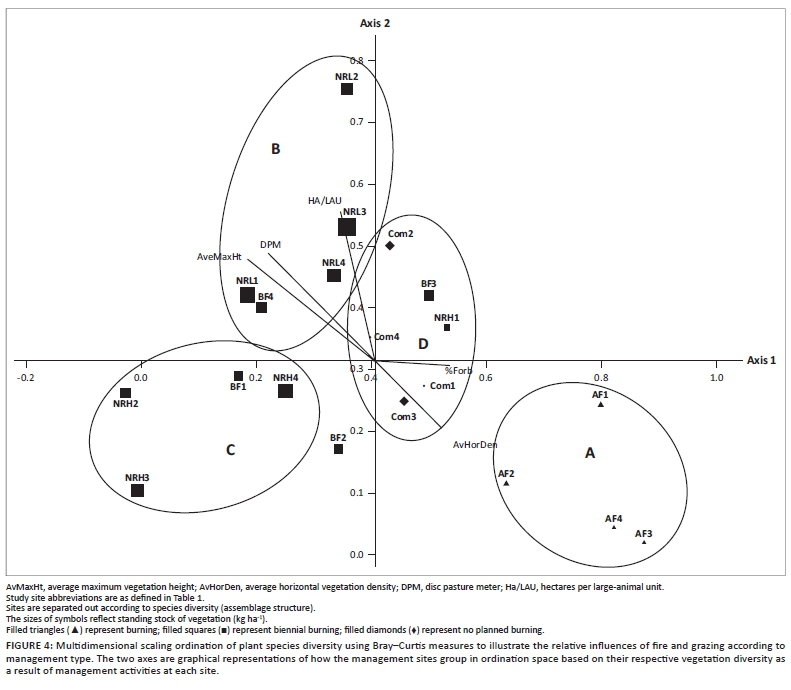
Grasses and forbs responded in a similar manner to management according to Mantel's asymptotic approximation (Mantel test, r = 0.26, P < 0.0005). The heavily grazed conservation area (NRH) showed significantly reduced abundance of decreaser grasses relative to increaser grasses (Z = -1.59, P < 0.05; Figure 3a) and had a high proportion of increaser III grasses. There was also a significantly lower overall abundance of palatable versus unpalatable grasses (Z = 1.76, P < 0.05; Figure 3b). The part of the nature reserve experiencing low grazing pressure supported the highest proportion of palatable grass species (Figure 4).

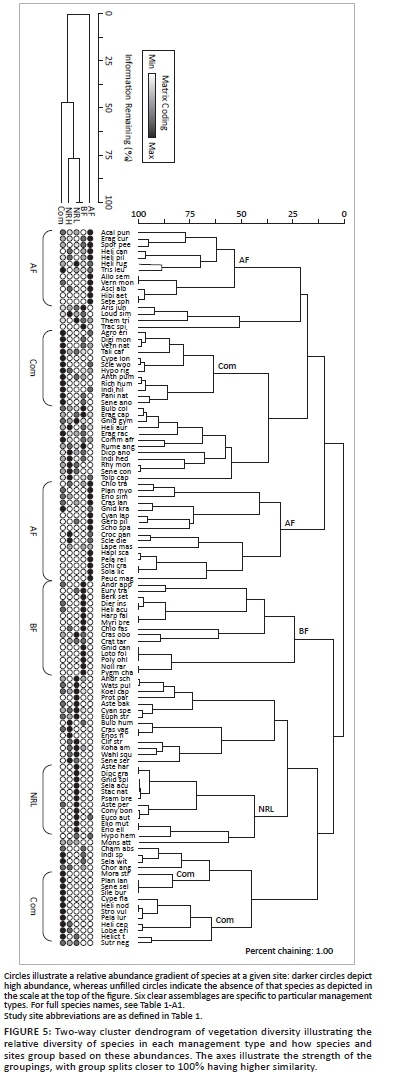
Fire frequency had an overriding effect on plant species diversity, with grazing intensity having a secondary role in distinguishing the effect of different management practices (Figure 4). Axis 1 separates sites according to burn frequency, whereas axis 2 indicates a response to grazing pressure. Sites in group B are defined primarily by the vegetation density as measured by DPM, average maximum vegetation height and stocking density. Sites in group A were mostly characterised by their average horizontal vegetation density (AvHorDen). Sites in group C were different from the rest but did not differ based on any specific parameter. Similarly, sites in group D were not found to be different from other sites based on any specific parameters, but shared aspects of each of the other groupings with regard to their vegetation parameters. Communal lands were defoliated to such an extent that it was difficult to distinguish the two responses. DPM data were overlaid to illustrate the importance of phytomass in separating out sites. Axis 1 extracted 29.8% of the original distance matrix and axis 2 extracted 19.9% (total 49.7%).
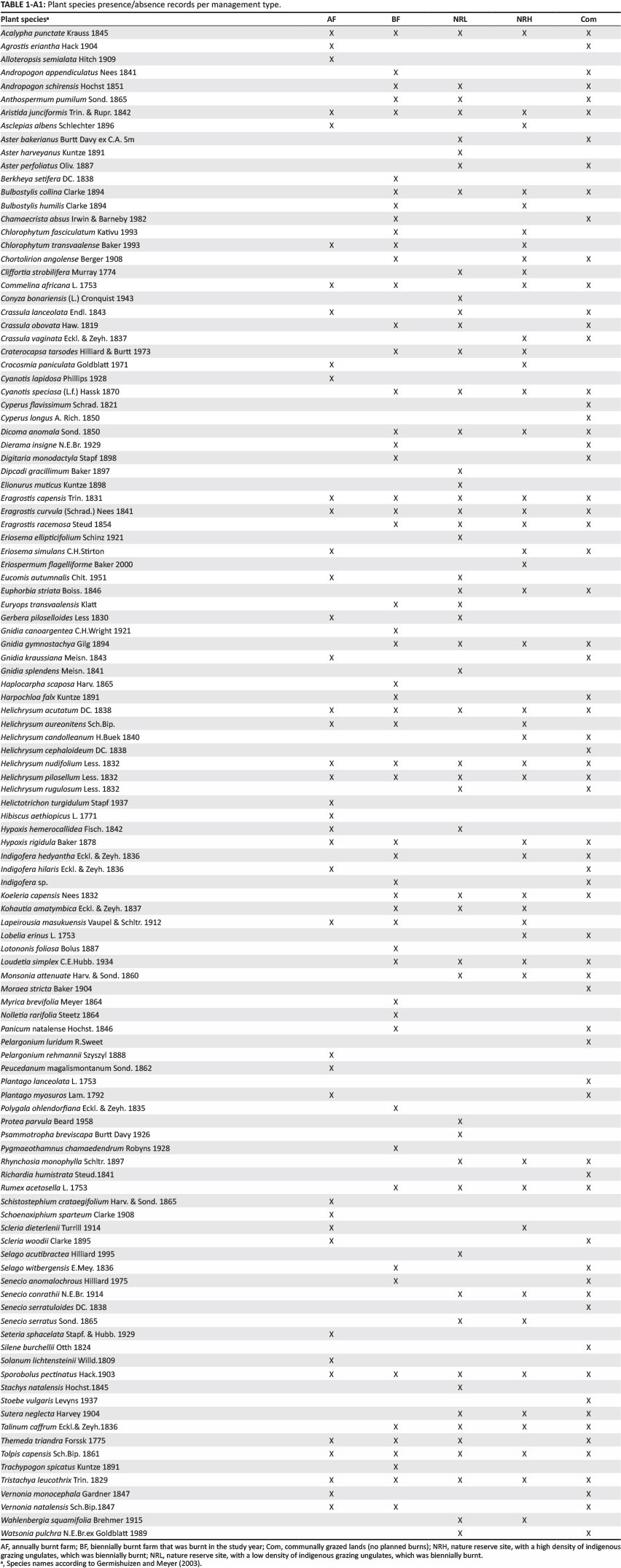
Plant species diversity responded strongly to disturbance (Figure 5). AF separates out first, followed by the sites Com, BF and NRL, which cluster very closely. These, in turn, cluster with NRH. Six distinct assemblages separate out according to land use. One of these is specific to the conserved area whereas the other five are associated with disturbed areas.
Discussion
Fire and grazing combined act as the primary disturbance mechanisms in grassland systems, shaping the structure and composition of the vegetation (Van Wilgen & Scholes1997; Veen et al. 2008). An understanding of the interplay between these disturbance effects in driving the system facilitates their being managed to influence ecosystem stability and biological intactness (Ferwerda et al. 2006). Fuhlendorf et al. (2012) accordingly suggested that fire and grazing should be viewed as essential ecosystem processes rather than tools for production goals. Annual burning (irrespective of litter accumulation), together with high grazing pressure (Everson 1999; Van Wilgen & Scholes 1997), has a detrimental effect on plant diversity in South Africa's moist highland grasslands (Joubert, Pryke & Samways 2014) and the high proportion of grasslands that are burnt annually is of concern for grassland conservation (Everson 1999).
Impact of management on vegetation structure
The current dominant management system in these grasslands combines high stocking rates (exceeding the ecological carrying capacity) with frequent (annual) fires.
To understand the effects of burning and grazing on vegetation structure, and to allow farmers to assess pasture condition rapidly, an easy-to-implement and efficient technique is required. The DPM has been recommended for these purposes (Bransby & Tainton 1977; Danckwerts & Trollope 1980; Trollope & Potgieter 1986). However, for this technique to be useful, the DPM must be calibrated for the vegetation type in question. The calibration for moist highland grasslands was both strong and linear and has a similar trend to previous calibrations in other grassland types.
Of the ten vegetation structure measures calculated in this study, four proved useful for illustrating the effects of different management practices (Table 2). Of these, three differentiated the impacts of management practices on vegetation structure. Patchiness was highest and vegetation cover (auto-correlated with grass cover because grass accounts for > 90% of vegetation cover) was lowest in the nature reserve stocked with ungulates (NHR). Although historically indigenous game species would not have grazed the high-altitude grasslands during the dry winter months (Mucina & Rutherford 2006:357), current management on a portion of the conservation area has indigenous ungulates fenced at relatively high density (13.5 ha LAU−1 - 14.5 ha LAU−1) throughout the year. Resultant selective grazing, predominantly by blesbok, leads to a high proportion of bare ground and unpalatable vegetation. Grazing herbivores modify the structural heterogeneity of vegetation within a particular successional sere (Milchunas et al. 1988; Owen-Smith & Danckwerts 1997), but vegetation biomass alone does not reflect this because biomass of the unpalatable component remains relatively high (Figures 2a and 3b). The data presented here suggest that indigenous game species, stocked at a high density, have a greater impact on both plant diversity and structure than domestic livestock. Even at fairly low stocking rates, selective grazing by indigenous game species clears all decreaser species and creates a mosaic of patchy, unpalatable vegetation (Figure 3). Few studies focusing on the potential impact of indigenous game species on moist highland grasslands have been carried out. Modern fencing practices for conservation, wildlife ranching or agriculture have restricted free movement of indigenous game species and, in some cases, resulted in overabundance and overgrazing in wildlife preserves (Coughenour 1991). Thus far, ecological studies in grassland systems have stressed the importance of fire more than native ungulate grazing (Wilsey & Martin 2015), which may be equally important when examining the ecological integrity of grassland systems.
By contrast, moderate to heavy bulk grazing by domestic livestock has no significant impacts on vegetation structure or diversity, provided that the burning interval is at least 2 years. However, the similarity of the AF and Com sites, which are burnt annually or have never been burnt intentionally, respectively, suggests that when grazing pressure is intense, the relative importance of fire in controlling vegetation structure diminishes. Few grassland specialist plant species require low disturbance levels and few pioneer species require high disturbance levels to thrive (Fox 1979).
In the study area, farmers introduce livestock immediately following a spring burn. As plant growth is limited to the summer period (Tainton, Groves & Nash 1977), this inevitably influences the phenological stages of grasses and eventually the vegetation structure. Excessive bulk grazing does not necessarily lead to loss of grass species (Figure 3b), because the basal meristem of grass leaves enables regrowth after defoliation. However, frequent and excessive defoliation can shift species assemblages towards grasslands being dominated by increaser II species (Milchunas et al. 1988; Figure 3a). As a result, horizontal vegetation density can be used as a surrogate for the presence of a 'carpet-like' layer of thick, low vegetation close to the ground. This habitat homogenisation is expected when grazing pressure is high and largely unselective (Swengel 2001). Not surprisingly, these conditions result in low vegetation biomass (Figure 1) and a correspondingly low carrying capacity for grazers (Figure 2a). Limb et al. (2011) illustrated that management that involves infrequent fire and moderate stocking rates results in improved livestock production.
Conservative management (e.g. biennial burning as seen in BF) resulted in high vegetation biomass, along with a greater proportion of decreaser grass species and an increase in overall palatability of the vegetation (Figures 2b and 3a, Table 5). When heavy grazing and frequent burning are combined, high horizontal vegetation density results. The nature reserve with low grazing levels (NRL) had the lowest horizontal vegetation density and thus current management practices in this part of the conserved area appear to be ecologically sound.
We recommend that a DPM method is used in future assessments of vegetation structure integrity and forage availability, without the need for any other form of sampling. The DPM (Figure 1) has proved an effective surrogate for all structural indices. By combining these data with grazing intensity (expressed as ha LAU−1), an estimate of forage availability can be obtained per LAU (referred to as a 'fodder capacity index') (Figure 2a). This new approach encompasses both the current standing stock of vegetation (which has already been affected by grazing and fire) and the future potential grazing impact based on current stocking rate. The approach recommended here is user friendly, time efficient and accurate, and allows for adaptive management.
The available stocks of forage on communal lands and the annually burnt farm are, on average, less than 1100 kg LAU−1 throughout the summer season. According to the recommended minimum of 3 ha LAU−1 (Tainton 1999), and assuming that the stocking rate on BF is sufficient for sustainability of current phytomass, the recommended minimum sustainable threshold of forage per LAU for domestic livestock should be 5000 kg LAU−1 (see Figure 2a, where BF standing stocks resemble those of NRL).
Impact of management on plant species diversity
The use of plant species as indicators of disturbance has been shown to be ineffective (Lindenmayer et al. 2002; Öster, Persson & Eriksson 2008); however, assemblages of grassland species reflect different disturbance pressures (McIntyre & Lavorel 1994; Milchunas et al. 1988). In our analysis, grasses and forbs responded in a similar manner to management, suggesting that either group on its own would be sufficient for assessment of plant ecological integrity. The presence of P. myosuros at AF is a strong signal of habitat degradation, as this species is a known weed and is often spread through livestock dung. Land degradation is characterised more by the gain of pioneer-type species than by the loss of specialist grassland species. The combination of heavy grazing and annual burning leads to a distinct plant community (group A in Figure 4), with nine species characterising this group. The large number of 'disturbance specialists' (species found only in the heavily disturbed AF, Com and NRH sites) illustrates the sensitivity of this system to assemblage shifts favouring increaser or pioneer species. High-intensity selective grazing by indigenous herbivores promotes a community of unpalatable species (Figure 3), which are largely generalist taxa (Figure 5). Of particular interest is the significantly reduced abundance of decreaser grasses relative to increaser grasses in the heavily grazed conservation area (Figure 3a). This is supported by the high proportion of increaser III grasses.
In Figure 4, group B (biennially burnt nature reserve with low grazing pressure) and group A (annually burnt farm) differ most markedly, as they are furthest apart in ordination space. This illustrates the differing effects of high-frequency burning (A) and low-intensity grazing (B). The low-intensity grazing within the nature reserve shows a floral community that is significantly different from all other sites (Table 4). The (rarely burnt) communal grazing lands (group D) and the biennially burnt lands are intermediate in plant community structure and do not separate out as clear groups. Grazing intensity and fire frequency are therefore considered complementary disturbance factors. The separation of sites is driven primarily by high phytomass associated with, in this case, low grazing intensity on the low disturbance/defoliation extreme and by high horizontal vegetation density and forb cover on the high disturbance/defoliation extreme.
The 114 plant species identified fall into six distinct management-specific communities and four generalist groupings (Figure 5), which implies that plants show strong responses to both grazing and burning. Two of the 114 species - E. autumnalis and H. hemerocallidea - are classified as threatened and both are listed as decreasing (Raimondo et al. 2009). Eucomis autumnalis, a popular plant for use in traditional medicine, is confined to the conserved area, suggesting that it is sensitive to disturbance.Hypoxis hemerocallidea, also used in traditional medicine, is confined to unconserved, communal lands that burn infrequently. The only detectable effect in areas grazed by domestic livestock and that are burnt biennially was the low representation of forbs (Scott-Shaw & Morris 2014). This suggests that grazing may have an important role in determining forb diversity; indeed, Uys et al. (2004) found that annual versus biennial burning made little difference to forb diversity in grassland systems. This may also be attributed to an intermediate disturbance effect, causing species to be lost both as a result of intensive management and in response to a lack of fire management when grasses become moribund, thereby stifling forbs; however, this would need to be illustrated more robustly.
Conclusion
The data presented here suggest that indigenous game species, stocked at high density, have a greater impact on both plant diversity and vegetation structure than domestic livestock. Even at fairly low stocking rates, selective grazing by indigenous game species clears all decreaser species and creates a mosaic of patchy, unpalatable vegetation. However, burning biennially (even with commercially viable stocking rates) resulted in high vegetation biomass along with a greater proportion of decreaser grass species and an increase in overall palatability of the vegetation.
Based on the results of this study, we recommend that future assessments of vegetation structure integrity and forage availability use a DPM method. Based on the novel 'fodder capacity index' presented here, the recommended minimum sustainable threshold of forage per LAU for high domestic livestock production and for sustaining biological diversity (and ecosystem services) should be 5000 kg LAU−1.
Acknowledgements
We thank Balongile Bhengu and her staff at Verloren Valei Nature Reserve for their support and assistance with historical data. Farm managers and owners (Thys, the Grobbelaars, Bafana and Job of Sakhelwe, and Roger Croall) are thanked for allowing the research to be conducted on their properties. We gratefully acknowledge Luciano Makaka, Maxwell Boakye, Thabo Mabuza, Thomas Birch, Jessica da Silva, Grant Cannon and others who assisted in the field. We are also grateful to Colin Everson for the data he provided and Thomas Birch for computer assistance. This research was funded by the National Research Foundation of South Africa, the South African National Biodiversity Institute, the Rufford Small Grant Foundation, Tshwane University of Technology and the University of Cape Town.
Competing interests
The authors declare that they have no financial or personal relationships that may have inappropriately influenced them in writing this article.
Authors' contributions
I.T.L. (University of Cape Town) was the project leader and was responsible for data collection, analysis and interpretation, as well as drafting the initial manuscript and raising funds for the research. P.A.R.H. (University of Cape Town) and R.J. (Tshwane University of Technology) were both responsible for project design and contributed to the analysis and interpretation of data, revising the manuscript and obtaining funding.
References
Allan, D.G., Harrison, J.A., Navarro, R.A., Van Wilgen, B.W. & Thompson, M.W., 1997, 'The impact of commercial afforestation on bird populations in Mpumalanga Province, South Africa-Insights from bird-atlas data', Biological Conservation 79, 173-185. http://dx.doi.org/10.1016/S0006-3207(96)00098-5 [ Links ]
Anderson, M.J., 2001, 'A new method for non-parametric multivariate analysis of variance', Austral Ecology 26, 32-46. http://dx.doi.org/10.1111/j.1442-9993.2001.01070.pp.x [ Links ]
Bond, W.J., 1997, 'Fire', in R.M. Cowling, D.M. Richardson & S.M. Pierce (eds.), Vegetation of Southern Africa, pp. 421-446, Cambridge University Press, Cambridge. [ Links ]
Bransby, D.I. & Tainton, N.M., 1977, 'The disc pasture meter: Possible applications in grazing management', Proclamations of the Grassland Society of South Africa 12, 115-118. http://dx.doi.org/10.1080/00725560.1977.9648818 [ Links ]
Bray, J.R. & Curtis, J.T., 1957, 'An ordination of the upland forest communities of Southern Wisconsin', Ecological Monographs 27, 325-349.http://dx.doi.org/10.2307/1942268 [ Links ]
Burgoyne, B.M., Bredenkamp, G.J. & Van Rooyen, N., 2000, 'Wetland vegetation in the North-Eastern Sandy Highveld, Mpumalanga, South Africa', Bothalia 30, 187-200.http://dx.doi.org/10.4102/abc.v30i2.558 [ Links ]
Carbutt, C., Tau, M., Stevens, A. & Escott, B., 2011, 'The conservation status of temperate grasslands in Southern Africa', Grassroots 11, 17-23. [ Links ]
Collins, S.L. & Calabrese, L.B., 2012, 'Effects of fire, grazing and topographic variation on vegetation structure in tallgrass prairie', Journal of Vegetation Science 23(3), 563-575. http://dx.doi.org/10.1111/j.1654-1103.2011.01369.x [ Links ]
Coughenour, M.B., 1991, 'Spatial components of plant-herbivore interactions in pastoral, ranching, and native ungulate ecosystems', Journal of Range Management 44(6), 530-542. [ Links ]
Danckwerts, J.E. & Trollope, W.S.W., 1980, 'Assessment of the disc pasture meter on natural veld in the false thornveld of the eastern province', Proceedings of the Annual Congresses of the Grassland Society of Southern Africa 15, 47-52. http://dx.doi.org/10.1080/00725560.1980.9648884 [ Links ]
Department of Environmental Affairs, 2011, National list of ecosystems that are threatened and in need of protection (notice 1002 of 2011), Government Gazette 34809, 9 December, Government Printer, Pretoria. [ Links ]
Douglas, M.E. & Endler, J.A., 1982, 'Quantitative matrix comparisons in ecological and evolutionary investigations', Journal of Theoretical Ecology 99, 777-795.http://dx.doi.org/10.1016/0022-5193(82)90197-7 [ Links ]
Dufrêne, M. & Legendre, P., 1997, 'Species assemblages and indicator species: The need for a flexible asymmetrical approach', Ecological Monographs 67, 345-366.http://dx.doi.org/10.2307/2963459 [ Links ]
Everson, C.S., 1999, 'Veld burning in different vegetation types: Grassveld', in N.M. Tainton (ed.), Veld management in South Africa, pp. 228-235, University of Natal Press, Pietermaritzburg. [ Links ]
Ferwerda, J.G., Siderius, W., Van Wieren, S.E., Grant, C.C., Peel, M., Skidmore, A.K. et al., 2006, 'Parent material and fire as principle drivers of foliage quality in woody plants', Forest Ecology and Management 231, 178-183. http://dx.doi.org/10.1016/j.foreco.2006.05.044 [ Links ]
Fox, J.F., 1979, 'Intermediate-disturbance hypothesis', Science 204, 1344-1345. http://dx.doi.org/10.1126/science.204.4399.1344 [ Links ]
Fuhlendorf, S.D., Engle, D.M., Elmore, R.D., Limb, R.F. & Bidwell, T.G., 2012, 'Conservation of pattern and process: Developing an alternative paradigm of rangeland management', Rangeland Ecology & Management 65, 579-589. http://dx.doi.org/10.2111/REM-D-11-00109.1 [ Links ]
Ganguli, A.C., Vermeire, L.T., Mitchell, R.B. & Wallace, M.C., 2000, 'Comparison of four nondestructive techniques for estimating standing crop in shortgrass plains', Journal of Agronomics 92, 1211-1215. http://dx.doi.org/10.2134/agronj2000.9261211x [ Links ]
Germishuizen, G. & Meyer, N.L., 2003, Plants of Southern Africa: An annotated checklist, South African National Biodiversity Institute, Pretoria (Strelitzia no. 14). [ Links ]
Heyns, J., 1985, Verslag oor die veldtoestand en weidingskapasiteit van die Steenkampsberg Naturreservaat [Report on the veld condition and grazing capacity of the Steenkampsberg Nature Reserve], [ Links ] report, Departement van Landbou en Watervoorsiening, Pretoria.
Hurlbert, S.H., 1984, 'Pseudoreplication and the design of ecological field experiments', Ecological Monographs 54, 187-211. http://dx.doi.org/10.2307/1942661 [ Links ]
Jacobs, B.F., Kingston, J.D. & Jacobs, L.L., 1999, 'The origin of grass-dominated ecosystems', Annals of the Missouri Botanical Garden 86(2), 590-643.http://dx.doi.org/10.2307/2666186 [ Links ]
Joubert, L., Pryke, J.S. & Samways, M.J., 2014, 'Annual burning drives plant communities in remnant grassland ecological networks in an afforested landscape', South African Journal of Botany 92, 126-133. http://dx.doi.org/10.1016/j.sajb.2014.02.009 [ Links ]
Le Maitre, D.C., Van Wilgen, B.W., Chapman, R.A. & McKelly, D.H., 1996, 'Invasive plants and water resources in the Western Cape Province, South Africa: Modelling the consequences of a lack of management', Journal of Applied Ecology 33, 161-172. http://dx.doi.org/10.2307/2405025 [ Links ]
Limb, R.F., Fuhlendorf, S.D., Engle, D.M., Weir, J.R., Elmore, R.D. & Bidwell, T.G., 2011, 'Pyric-herbivory and cattle performance in grassland ecosystems', Rangeland Ecology & Management 64, 659-663. http://dx.doi.org/10.2111/REM-D-10-00192.1 [ Links ]
Lindenmayer, D.B., Cunningham, R.B., Donnelly, C.F. & Lesslie, R., 2002, 'On the use of landscape surrogates as ecological indicators in fragmented forests', Forest Ecology and Management 159, 203-216. http://dx.doi.org/10.1016/S0378-1127(01)00433-9 [ Links ]
Lipsey, M.K. & Hockey, P.A.R., 2010, 'Do ecological networks in South African commercial forests benefit grassland birds? A case study of a pine plantation in KwaZulu-Natal', Agriculture Ecosystem and Environment 137, 133-142. http://dx.doi.org/10.1016/j.agee.2010.01.013 [ Links ]
Little, I.T., 2011, 'Bird reproductive success and faunal habitat selection as tools for understanding the impacts of land-use management on moist highland grassland biodiversity in South Africa', PhD thesis, Dept. of Zoology, University of Cape Town. [ Links ]
Little, I.T., Hockey, P.A.R. & Jansen, R., 2013, 'A burning issue: Fire overrides grazing as a disturbance driver for South African grassland bird and arthropod assemblage structure and diversity', Biological Conservation 158, 258-270. http://dx.doi.org/10.1016/j.biocon.2012.09.017 [ Links ]
Little, I.T., Little, R.M., Jansen, R. & Crowe, T.M., 2005, 'Winter bird assemblages, species richness and relative abundance at a re-vegetated coal mine in the Middelburg district, Mpumalanga province, South Africa', South African Journal of Wildlife Research 35, 13-22. [ Links ]
MacDonald, I.A.W., 1989, 'Man's role in changing the face of Southern Africa', in B.J. Huntley (ed.), Biotic diversity in Southern Africa - Concepts and conservation, pp. 51-77, Oxford University Press, Cape Town. [ Links ]
Manry, D.E. & Knight, R.S., 1986, 'Lightning density and burning frequency in South African vegetation', Vegetatio 66, 67-76. http://dx.doi.org/10.1007/BF00045496 [ Links ]
McCune, B. & Grace, J.B., 2002, Analysis of ecological communities, MjM Software Design, Gleneden Beach. [ Links ]
McCune, B. & Mefford, M.J., 2006, PC-ORD: Multivariate analysis of ecological data, MjM Software Design, Version 5.10., Gleneden Beach. [ Links ]
McIntyre, S. & Lavorel, S., 1994, 'How environmental and disturbance factors influence species composition in temperate Australian grasslands', Journal of Vegetation Science 5, 373-384. http://dx.doi.org/10.2307/3235861 [ Links ]
McNaughton, S.J., 1986, 'Grazing lawns: On domesticated and wild grazers', American Naturalist 128, 937-939. [ Links ]
Mentis, M.T., 1981, 'The effect of scale of burn on the densities of grassland francolins in the Natal Drakensberg', Biological Conservation 21, 247-261.http://dx.doi.org/10.1016/0006-3207(81)90081-1 [ Links ]
Milchunas, D.G., Sala, O.E. & Lauenroth, W.K., 1988, 'A generalized model of the effects of grazing by large herbivores on grassland community structure', American Naturalist 132, 87-106. [ Links ]
Mucina, L. & Rutherford, M.C. (eds.), 2006, The vegetation of South Africa, Lesotho and Swaziland, South African National Biodiversity Institute, Pretoria (Strelitzia no. 19). [ Links ]
Neke, K.S. & Du Plessis, M.A., 2004, 'The threat of transformation: Quantifying the vulnerability of grasslands in South Africa', Conservation Biology 18, 466-477.http://dx.doi.org/10.1111/j.1523-1739.2004.00157.x [ Links ]
O'Connor, T.G., 2005, 'Influence of land use on plant community composition and diversity in Highland Sourveld grassland in the southern Drakensberg, South Africa',Journal of Applied Ecology 42, 975-988. http://dx.doi.org/10.1111/j.1365-2664.2005.01065.x [ Links ]
O'Connor, T.G., Kuyler, P., Kirkman, K.P. & Corcoran, B., 2010, 'Which grazing management practices are most appropriate for maintaining biodiversity in South African grassland?', African Journal of Range & Forage Science 27, 67-76. http://dx.doi.org/10.2989/10220119.2010.502646 [ Links ]
O'Connor, T.G., Martindale, G., Morris, C.D., Short, A., Witkowski, E.T.F. & Scott-Shaw, R., 2011, 'Influence of grazing management on plant diversity of Highland Sourveld grassland, KwaZulu-Natal, South Africa', Rangeland Ecology and Management 64, 196-207. http://dx.doi.org/10.2111/REM-D-10-00062.1 [ Links ]
Öster, M., Persson, K. & Eriksson, O., 2008, 'Validation of plant diversity indicators in semi-natural grasslands', Agriculture Ecosystem and Environment 125, 65-72.http://dx.doi.org/10.1016/j.agee.2007.11.006 [ Links ]
Owen-Smith, N., 1999, 'The animal factor in veld management', in N.M. Tainton (ed.), Veld management in South Africa, pp. 117-138, University of Natal Press, Pietermaritzburg. [ Links ]
Owen-Smith, N. & Danckwerts, J.E., 1997, 'Herbivory', in R.M. Cowling, D.M. Richardson & S.M. Pierce (eds.), Vegetation of Southern Africa, pp. 397-420, Cambridge University Press, Cambridge. [ Links ]
Raimondo, D., Van Staden, L., Foden, W., Victor, J.E., Helme, N.A., Turner, R.C. et al., 2009, Red list of South African plants 2009, South African National Biodiversity Institute, Pretoria (Strelitzia no. 25). [ Links ]
Rowe-Rowe, D.T. & Scotcher, J.S.B., 1986, 'Ecological carrying capacity of the Natal Drakensberg for wild ungulates', South African Journal of Wildlife Research 16, 12-16. [ Links ]
Scott-Shaw, R. & Morris, C.D., 2014, 'Grazing depletes forb species diversity in the mesic grasslands of KwaZulu-Natal, South Africa', African Journal of Range and Forage Science 32, 21-31. http://dx.doi.org/10.2989/10220119.2014.901418 [ Links ]
Smith, B., 2006, The farming handbook, Interpak Books, Pietermaritzburg. [ Links ]
South African National Biodiversity Institute, 2014, Grazing and burning guidelines: Managing grasslands for biodiversity and livestock production, South African National Biodiversity Institute, Pretoria. [ Links ]
Sternberg, M., Gutman, M., Perevolotsky, A., Ungar, E.D. & Kigel, J., 2000, 'Vegetation response to grazing management in a Mediterranean herbaceous community: A functional group approach', Journal of Applied Ecology 37, 224-237. http://dx.doi.org/10.1046/j.1365-2664.2000.00491.x [ Links ]
Stohlgren, T.J., Falkner, M.B. & Schell, L.D., 1995, 'A modified-Whittaker nested vegetation sampling method', Vegetatio 117, 113-121.http://dx.doi.org/10.1007/BF00045503 [ Links ]
Swengel, A.B., 2001, 'A literature review of insect responses to fire, compared to other conservation managements of open habitat', Biodiversity and Conservation 10, 1141-1169. http://dx.doi.org/10.1023/A:1016683807033 [ Links ]
Tainton, N.M., 1999, Veld management in South Africa, University of Natal Press, Pietermaritzburg. [ Links ]
Tainton, N.M., Groves, R.H. & Nash, R., 1977, 'Time of mowing and burning veld: Short term effects on production and tiller development', Proceedings of the Annual Congresses of the Grassland Society of Southern Africa 12, 59-64. http://dx.doi.org/10.1080/00725560.1977.9648806 [ Links ]
Trollope, W.S.W. & Potgieter, A.L.F., 1986, 'Estimating grass fuel loads with a disc pasture meter in the Kruger National Park', Journal of the Grassland Society of Southern Africa 3, 148-152. http://dx.doi.org/10.1080/02566702.1986.9648053 [ Links ]
Uys, R.G., Bond, W.J. & Everson, T.M., 2004, 'The effect of different fire regimes on plant diversity in southern African grasslands', Biological Conservation 118, 489-499.http://dx.doi.org/10.1016/j.biocon.2003.09.024 [ Links ]
Van Rooyen, N., Bredenkamp, G.J. & Theron, G.K., 1986, 'Veld management', in J.d.P. Bothma (ed.), Game ranch management, pp. 539-578, Van Schaik, Pretoria. [ Links ]
Van Wilgen, B.W. & Scholes, R.J., 1997, 'The vegetation and fire regimes of southern hemisphere Africa', in B.W. van Wilgen, M.O. Andreae, J.G. Goldammer & J.A. Lindesay (eds.), Fire in Southern African savannas: Ecological and atmospheric perspectives, pp. 27-46, Witwatersrand University Press, Johannesburg. [ Links ]
Veen, G.F., Blair, J.M., Smith, M.D. & Collins, S.L., 2008, 'Influence of grazing and fire frequency on small-scale plant community structure and resource variability in native tallgrass prairie', Oikos 117, 859-866. http://dx.doi.org/10.1111/j.0030-1299.2008.16515.x [ Links ]
Wiens, J.A. & Rotenberry, J.T., 1981, 'Habitat associations and community structure of birds in shrubsteppe environments', Ecological Monographs 51, 21-41.http://dx.doi.org/10.2307/2937305 [ Links ]
Wilsey, B.J. & Martin, L.M., 2015, 'Top-down control of rare species abundances by native ungulates in a grassland restoration', Restoration Ecology 23, 465-472.http://dx.doi.org/10.1111/rec.12197 [ Links ]
 Correspondence:
Correspondence:
Ian Little
Endangered Wildlife Trust, 3 Sangster Road, Howick 3290
South Africa
ianl@ewt.org.za
Received: 07 Dec. 2014
Accepted: 12 Aug. 2015
Published: 26 Nov. 2015
†, 1956-2013. This article is based on the thesis 'Bird reproductive success and faunal habitat selection as tools for understanding the impacts of land-use management on moist highland grassland biodiversity in South Africa' for the degree of Doctor of Philosophy in the Percy FitzPatrick Institute of African Ornithology, Department of Zoology, University of Cape Town (July 2011). Supervised by Prof. Philip Hockey and co-supervised by Prof. Raymond Jansen.
Appendix 1


{kind=link}