










Services on Demand
Journal

Article
 English (pdf)
English (pdf)
 Article in xml format
Article in xml format Article references
Article references
 Send this article by e-mail
Send this article by e-mailIndicators
Related links
-
 Cited by Google
Cited by Google -
 Similars in Google
Similars in Google
Share

 Permalink
PermalinkSouth African Journal of Science
On-line version ISSN 1996-7489Print version ISSN 0038-2353
S. Afr. j. sci. vol.116 n.3-4 Pretoria Mar./Apr. 2020
https://doi.org/10.17159/sajs.2020/6542
RESEARCH ARTICLE
Pleistocene large reptile tracks and probable swim traces on South Africa's Cape south coast
Charles W. HelmI, II; Hayley C. CawthraI, III; Xander CombrinkIV; Carina J.Z. HelmV; Renée RustVI; Willo StearI; Alex van den HeeverVII
IAfrican Centre for Coastal Palaeoscience, Nelson Mandela University, Port Elizabeth, South Africa
IIPeace Region Palaeontology Research Centre, Tumbler Ridge, British Columbia, Canada
IIIGeophysics and Remote Sensing Unit, Council for Geoscience, Cape Town, South Africa
IVDepartment of Nature Conservation, Tshwane University of Technology, Pretoria, South Africa
VDepartment of Geography, University of British Columbia, Vancouver, British Columbia, Canada
VISchool of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, Johannesburg, South Africa
VIITracker Academy, Magazine Hill, Graaff-Reinet, South Africa
ABSTRACT
The Cape south coast of South Africa contains a wealth of Pleistocene vertebrate trace fossil sites in aeolianites and cemented foreshore deposits. Published studies have described mammal and avian tracksites identified along this coastline. We report here on a number of Pleistocene palaeosurfaces within the Garden Route National Park that exhibit tracks of large reptiles, including probable swim traces. The tracks were probably made by more than one species, and may include a crocodylian. There are no extant reptiles in this coastal region capable of making such tracks and traces, which probably represent an indication of a previously more extensive range for the Nile crocodile and a monitor lizard. These findings demonstrate the potential for ichnology to complement the traditional body fossil record. Two Middle Stone Age stone artifacts were found embedded in one palaeosurface containing multiple reptile trackways. These discoveries have implications for the understanding of Pleistocene palaeoenvironment and palaeoclimate - in an area which is important in the study of modern human origins.
SIGNIFICANCE:
• Large reptile Pleistocene fossil tracksites have recently been discovered on the Cape south coast of South Africa where there are no previous such records, and no reptiles of this size are currently found in the region.
• These sites include the first reported probable reptile swim traces in Africa and one tracksite also contained two Middle Stone Age artifacts.
• These discoveries have implications for Pleistocene environments and climate on the Cape south coast.
Keywords: Pleistocene reptiles, fossil tracks, swim traces, crocodylians, lithics
Introduction
Fossil tracks and trackways are common in Middle and Late Pleistocene coastal aeolianites and cemented foreshore deposits along the Cape south coast of South Africa.1-7 These Pleistocene rocks have been the focus of a 350-km ground survey by a team led by C.W.H. between 2007 and 2019, from Arniston in the west to Robberg in the east. The resulting corpus of publications has dealt with mammal tracks and avian tracks. The only mention of reptile tracks was by Roberts et al.1 who indicated that tracks of testudinae had been found in aeolianites east of Still Bay. However, no details were provided, and the site has not been re-located. Likewise, the fossil, archaeological and historical records have largely remained silent with regard to large terrestrial reptiles along this coastline, other than occasional reports of sea turtles, whereas there have been numerous reports on the mammalian fauna from archaeological sites, carnivore dens and scavenger dens.8-10
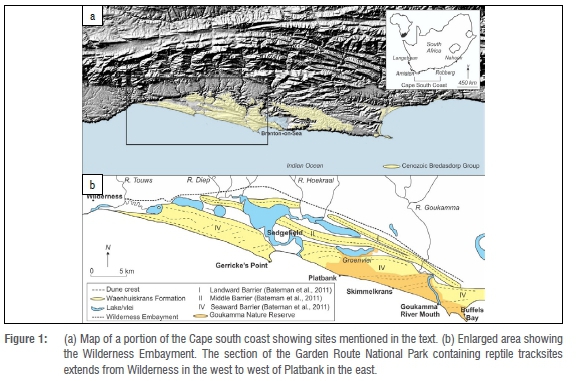
In 2018 our team identified palaeosurfaces containing tracks and trackways of large reptiles, and what we interpret as swim traces, in a coastal section of the Garden Route National Park (Figure 1). One of these surfaces also contained Middle Stone Age (MSA) lithics. The purpose of this article is to describe the tracks and track forms of these reptiles, consider plausible trackmakers, describe the lithics, and consider the palaeoenvironmental implications of these discoveries.
Geological context
Pleistocene dunes and beaches occur as aeolianites and cemented foreshore deposits along the Cape south coast.11-14 Aeolianite facies form the Waenhuiskrans Formation15, and foreshore facies form part of the Klein Brak Formation, which Malan16 interpreted as representing a succession of shallow marine, beach and estuarine or lagoonal deposits. These two formations form major elements of the Bredasdorp Group.15,16 One of Malan's16 stratotypes for the Klein Brak Formation is exposed at the Swartvlei estuary, situated east of the sites we describe.
The tracksites described lie within the Wilderness Embayment, the stratigraphic record of which has been described.11,17,18 Bateman et al.11 applied optically stimulated luminescence (OSL) dating to sites in this area: samples obtained from close to the tracksites we report on yielded a range of dates, the oldest being 148 ka ± 10 ka and the youngest being 92 ka ± 5 ka. Stratigraphy along this coastline consists mostly of composite Marine Isotope Stage (MIS) 6 and MIS 5 deposits, draped by a Holocene unconsolidated unit of dunes.11 Around 91 ka, sea levels were as much as 45 m lower than at present, and the coastline in this area may have been as much as 60 km seaward of today's coast.19 Around 126 ka, sea levels were 6.6-8 m higher than present levels on the Cape south coast.20 Exposure of the continental shelf during sea-level lowstands created an environment characterised by lakes, lagoons, rivers and estuaries.21
Although palaeo-dunes generally dominate the Cape south coast compared to more rarely preserved beach deposits12, Carr et al.20 mapped out evidence of the MIS 5e highstand at Cape Agulhas, the Great Brak River mouth and Swartvlei Estuary. Further work was conducted at the Great Brak River mouth by Cawthra et al.13, who described a regressive succession of shoreface, foreshore and aeolian deposits.
Methods
Once the tracks were identified in 2018, a search was conducted in the surrounding area for similar palaeosurfaces. Global Positioning Survey readings were recorded for all sites using a handheld device. Locality data were reposited with the African Centre for Coastal Palaeoscience and Garden Route National Park, to be made available to bona fide researchers upon request.
Measurements of tracks and traces included length, width, depth, pace length and stride length.22 Results were recorded in centimetres. Expert trackers were invited to comment on tracks and potential trackmakers, through photographs and site visits. Tracings were made of selected tracks. Photographs were taken for photogrammetric analysis.23 Three-dimensional models were generated with Agisoft MetaShape Professional (v. 1.0.4) using an Olympus TG-5 camera (focal length 4.5 mm; resolution 4000 x 3000; pixel size 1.56 x 1.56 um). The final images were rendered using CloudCompare (v.2.10-beta).
As no in-situ tracksites were evident, in order to estimate the original angle of the track-bearing surfaces, we considered the orientation of rhizoliths in underlying layers, the position and size of displacement rims, and the degree of track asymmetry. While the sites could be visited during most tidal conditions, low tide proved optimal. Tracksites were examined at different times of day, to allow sunlight to optimally illuminate the surfaces. Standard field techniques were applied in understanding tracksite context. Dip and strike measurements were taken on primary beds, bed thicknesses were measured, and textures were described.
A site visit was undertaken to evaluate the stone artifacts. Preliminary observations and descriptions were made. As the artifacts were embedded in the palaeosurface, not all their aspects could be examined.
Results
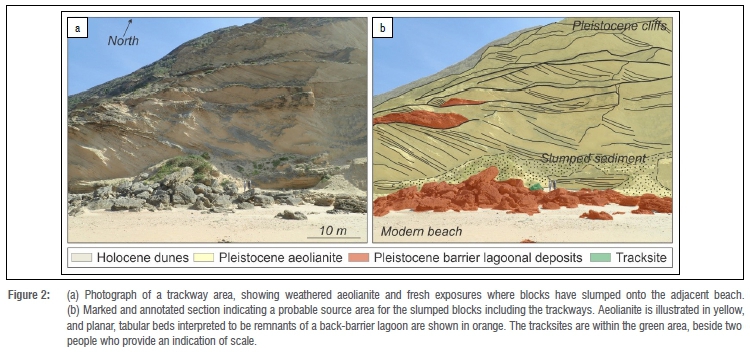
None of the tracksites we describe were in situ; all occur on loose blocks and slabs of varying size. We were not able to conclusively identify the levels of origin of these rocks, but field correlation suggested that they may be located close to their original locations, near the foot of the coastal cliffs at or near the level of planar-bedded deposits that have the same primary sedimentary structures, approximately 10 m above the modern beach (Figure 2). As the cliffs were unstable and safe access was not feasible, this could not be confirmed.
In total, 14 track-bearing blocks and slabs were identified on the eastern and western margins of a headland. The westernmost and easternmost track-bearing surfaces were ~330 m apart. Some tracksites were on small slabs, containing just a single track. More information could be gleaned from larger surfaces, which typically contained multiple tracks.
A range of track sizes was apparent, with the largest tracks measuring ~22 cm in length and the smallest ~10 cm in length. All tracks were on natural mould surfaces, with the tracks forming impressions in the surface (concave epirelief). In some cases we concluded that the original surface had been close to horizontal, while in other cases it appeared to have been at an angle of up to 30°. Here we first present results from four larger surfaces, which we named Surfaces A, B, C and D.
Regardless of locality, all tracksites were preserved in comparable stratigraphy and lithology. Fine- to medium-grained, well-sorted sand that exhibited planar beds up to 20 cm in thickness, alternated with trough-cross bedding, dominated this geological unit. The vertical orientation of fallen blocks could be determined by the presence of the tracks. The surface stratum consisted of a thin (less than 2 cm) layer of silt and fine sand. This veneer of finer-grained material was apparently laterally persistent as it was observed in association with tracks on both margins of the headland. The track-bearing layer inevitably occurred above and in contact with a heavily bioturbated layer up to 2 m in thickness. Rhizoliths occurred within these bioturbated sediments.
Surface A
The westernmost surface, which was inclined and east facing, measured 230 cm x 120 cm, on a loose rock 1.7 m in thickness (Figure 3a). Faint tracks could be discerned over much of the surface. Four large tracks were evident at the southern end (Figure 3b). Direction of motion was up the present slope of the surface. The two tracks on the left represented left-sided tracks, the two tracks on the right represented right-sided tracks, and the resulting trackway exhibited a wide straddle of 32 cm. The right upper (distal) track was orientated as much as 70° leftward compared to the other tracks. Substantial displacement rims were present posterolateral to the left proximal track and the right distal track.

The right proximal (lower) track was registered by the right forefoot. It was a partial track (being intersected by the bottom edge of the rock), and only showed the anterior ends of digital impressions.
The left proximal (lower) track was registered by the left hindfoot, and was characterised by a substantial displacement rim posterolaterally. Excluding the displacement rim but including the claw impressions, it was 22 cm long and 16 cm wide. Digit impressions were clearly evident; the longest was over 2.5 cm. Distance from the anterior edge of digits to the anterior end of the two claw impressions that were evident was 4 cm.
The left distal (upper) track was registered by the left forefoot, and was -18-19 cm long and 16 cm wide. It exhibited at least five digit impressions. Digits II, III and IV were each -7 cm long. Claw impressions were evident. The right distal (upper) track was made by the right hindfoot. It was less well preserved, but three deep digit depressions were evident, behind which were wide grooves, ending in a posterolateral displacement rim. Length could not be determined with precision; width was 16 cm.
The distance between hindfoot and forefoot (seen in the left tracks) was substantially less than the distance between forefoot and hindfoot (seen in the right tracks).
Surface B
Surface B lay 20 m east of Surface A, and was steeply inclined and west-facing. It measured 300 cm x 300 cm (Figure 4a). This surface presented a complex tracksite containing multiple reptile tracks and trackways, avian tracks, invertebrate burrow traces, and two stone artifacts.



The orientation of rhizoliths in the underlying bioturbated sediments suggested that the original surface had lain at an angle of -30° (Figure 4b). This supposition was supported by the asymmetrical nature of tracks that crossed the surface, with upslope margins being steeper than downslope margins. Three trackways crossed the surface transversely, and isolated tracks were orientated in an upslope direction. Some of these formed composite tracks. A zone of parting lineation of flute casts covered the upper right portion of the surface. This veneer obscured some areas in which extensions of trackways might have been registered.
Despite the cornucopia of tracks, trackway interpretation was challenging: one of the trackways appeared partially covered by the above-mentioned sediment veneer, two trackways appeared to exhibit tracks heading in both directions (resulting in composite tracks), track size and pace lengths were not consistent, and some tracks were poorly defined. Nonetheless, some individual tracks displayed well-preserved features.
One of the smaller depressions heading upslope was well preserved and showed what may have been four relatively parallel forward-pointing digits (Figure 4c). The posterior end tapered into a long, curved groove.
Deep digit impressions, similar to those seen on Surface A, and with a longer digit length (8 cm), were seen in one of the tracks heading upslope (Figure 4d). These were present at the distal end of a large, deep, composite impression. There were tantalising hints of parts of very large tracks at the bottom edge of the surface.
One of the tracks in the middle transverse trackway exhibited well-preserved narrow grooves that may represent claw scrape marks (Figure 4e). This trackway exhibited marked track asymmetry, with steep upslope margins and gentler downslope margins. The wide straddle typical of reptile tracks was not evident, and a reptile origin could therefore not be concluded.
In the upper transverse trackway, beside tracks heading in both directions, was a well-preserved smaller track, 17 cm long and 7 cm wide, with three small depressions (consistent with claw impressions) ahead of three of the digits (Figure 4f). The distance between the anterior ends of the digits and the anterior ends of the claw impressions ranged from 3 cm to 4 cm. In the upper right section, heading upslope, was a small trackway containing three tracks, with four digits in an anisodactyl pattern consistent with an avian trackmaker, with tracks 3.0 cm long and 3.2 cm wide, and pace length of 13-14 cm.
The two stone artifacts (Figure 5) were embedded in the upper end of the surface, right of centre. They occurred beside the zone of parallel parting plane lineations.

The artifact (flake) to the left in Figure 5a (and Figure 5b), appeared to be composed of silcrete or quartzite; if it was the former, then its reddish hue suggested that it may have been heat treated. It measured 5.5 cm x 3.5 cm. The ventral surface and left edge (as viewed in Figure 5a) were embedded in the matrix. The right edge appeared to have some retouch. The butt of the striking platform, the proximal end on which the detachment blow or pressure had been applied, appeared to have broken off, possibly as potential preparation for hafting. The remaining medial edge of this flake was cracked; this cracking may have been due to internal flaws of the material or from heating of the silcrete. Negative scars were visible on the dorsal surface.
The artifact (flake) to the right in Figure 5a was of milky quartz material with a distal end embedded in the matrix. It appeared that the dorsal surface was exposed. It measured 5.0 cm x 3.0 cm.
The flakes were probably struck off prepared cores, although the milky quartz flake may be considered a small core. There were no chips or debitage on the surface to suggest that the flakes had been produced at the site.
Early in 2020, a substantial landslide buried Surface B. A nearby (nonreptile) tracksite was also buried in this event.
Surface C
The easternmost large surface, west-facing, measured 220 cm in height and 200 cm in maximum width. Rhizoliths were evident, most prominently in the bottom right portion. Much of the middle section exhibited scour marks and flute casts caused by water flow (from left to right) over a sand surface. At the top end were three deep, large impressions: a single upper impression, and two impressions in a lower row (Figure 6a).
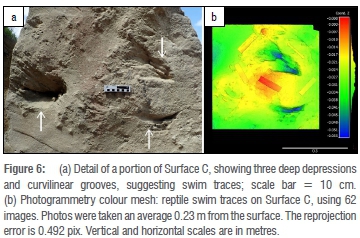
The two impressions in the lower row exhibited a characteristic, deep, curvilinear outline, 18 cm long, with convexity towards the bottom of the surface. To the left of each impression (as viewed in Figure 6a) were narrower curvilinear groove features, approximately 1 cm in diameter, ~2-3 cm apart, and 20-35 cm long (Figure 6b). These grooves extended through two of the larger depressions. If direction of movement was from right to left and the bottom row represented left tracks and the top impression was a right track, then these curvilinear grooves occurred anterior to the track impressions, and had a symmetrical appearance with outward convexity.
Surface D
Surface D lay 2 m to the west of Surface C. It was gently north-facing, measuring 215 cm x 60 cm (Figure 7a).


Surface D contained linear or curvilinear grooves, some of which were associated with digit impressions or larger, deeper impressions (Figure 7b, 7c).
The groove features were orientated in an approximately unimodal fashion, and were ~10 cm long. On a smaller surface beside Surface D were whitish cylindrical raised features (maximum dimensions 1.5 cm x 0.5 cm). Although rhizoliths and root casts occurred on nearby surfaces, and the shape of these features was not inconsistent with rhizoliths, macroscopically they appeared to contain osseous fragments. We therefore considered them more likely to be coprolites. They will be subjected to further analysis and will be reported on elsewhere.
Other sites
The remaining 10 sites confirmed that such tracks were a common local feature (Figure 8a-d); the majority of exposures of the distinctive track-bearing layer harboured reptile tracks. A small surface exposure situated between Surface A and Surface B contained a partial track with well-defined digit impressions (Figure 8a, 8b). We noted small reptile tracks (tortoise and perhaps terrapin) on two nearby surfaces with similar lithology. These will be described elsewhere.

Discussion
Track interpretation and trackmaker identification
The general crispness of the margins of the tracks is not consistent with a non-cohesive substrate such as soft, dry sand, and the depth of the tracks is not consistent with a very firm substrate.
The tracks appear to have been made on soft, wet substrates or underwater, in some cases on sloping surfaces, as suggested by the orientation of rhizoliths in underlying sediments. These factors may result in distortion of tracks, or sub-optimal preservation, compared with tracks made on firmer, drier surfaces. Suction effects may occur when a trackmaker's foot is removed from such a substrate, causing further distortion. This may explain the challenges we experienced in trackmaker identification despite the presence of many tracks. We interpreted the tracks of Surface A (Figure 3) as having been registered on a wet surface; the nature of the well-defined tracks, without evidence of swim traces, is not consistent with a deeper aqueous environment. This interpretation was buttressed by the presence of substantial displacement rims. The wide straddle, along with the distance between hindfoot and forefoot being less than that between forefoot and hindfoot, are consistent with the usual trackway pattern of a large reptile.22,24,25 The more leftward orientation of the right upper (distal) track suggested a substantial swivel to the left. This interpretation was supported by the possible surfeit of digit impressions in the left upper (distal) track, which probably represented removal of the left forefoot from the surface, followed by its re-placement on the surface with a more leftward bearing. Furthermore, we interpreted the posterolateral displacement rims as being consistent with forward propulsion and a swivel to the left.
Surface B (Figure 4) represented an inclined surface; with evidence of having been wet or submerged when the tracks were registered. While we did not find conclusive evidence of tail drag impressions, we note that such impressions are less likely to be evident when tracks are registered on submerged surfaces. The zone of parting lineation of flute casts was consistent with water flow from the upper section towards the lower section. The sandstone veneer containing these flute casts, which obscured some portions of trackways, was thus younger than the track-bearing layer.
We interpreted the tracks on Surface C (Figure 6) as probably having been made on a submerged surface, based in part on the presence of the scour marks and flute casts that indicated the presence of water flow. The presence of possible swim traces supported this interpretation. We also interpreted the tracks on Surface D (Figure 7) as having been registered on a submerged surface; in this case the appearance of the curvilinear groove features, connecting with digit impressions or deeper impressions, suggested a relatively shallow aqueous environment.
We have considered extant southern African reptiles that arguably could have created such tracks and trackways. These include (1) the Nile crocodile (Crocodylus niloticus), which belongs to the family Crocodylidae, and is the only extant crocodylian (order Crocodylia) in southern Africa,26 and (2) the two largest extant lizards in southern Africa, the water (Nile) monitor (Varanus niloticus) and rock monitor (Varanus albigularis), which belong to the order Squamata, family Varanidae.
During sea-level regressions in the terminal portion of the Middle Pleistocene and the Late Pleistocene, the vast Palaeo-Agulhas Plain was exposed along the Cape south coast.27 It was characterised by extensive floodplains and wetlands,19,28 and a distinctive megafauna of which a number of species are extinct. These wetlands may have provided habitat for large reptiles, although the warm conditions typically associated with large reptiles26 may not have been present. In assessing the track evidence, we also need to consider the possibility, albeit remote, that tracks may have been registered by an extinct, hitherto undescribed reptile.
The height of the possible source horizon is above present sea level; based on this height, as well as the chronology of the coastal cliff sequences,11 we posit an age of MIS 5e. Wetland environments, lagoons and back-barrier systems are well preserved in the highstand record as well as on the continental shelf, with their facies commonly identified in Pleistocene coastal cliffs.1,11,13,20 Our field evidence suggests that these trackways were mostly made in a sub-aqueous environment - interpreted to have been a back-barrier lagoon or interdune lake. The silt-rich stratum in which the tracks are preserved suggests depositional quiescence which, in coastal environments, is representative of a low-energy system such as a lagoon or interdune lake.29
The track and trackway morphology (digit shape, digit size, claw impressions, number of digits, wide straddle) provide evidence of large reptile trackmakers.22,24,25 The challenge is to determine whether these trackmakers can be identified to family or genus level, and whether or not more than one trackmaker species may have created the tracks. We do not believe that differences in track morphology between the two Varanus species are substantial enough to allow us to distinguish between them if they occur on these palaeosurfaces.24,25 However, V. albigularis would be a less likely trackmaker in an aquatic environment. The differences between the tracks of C. niloticus and V. niloticus are more substantial, including shape and orientation of the digits and distance between the anterior end of the toe impressions and the claw impressions (typically greater in Varanus).24,25
Using a formula devised by Thomson30 for estimating the length of C. niloticus based on the size of the hindfoot, and applying this to Surface A, a value of 257 cm was obtained. We are not aware of a similar formula for estimating length of Varanus species. We do not use track size as a sole criterion, because post-depositional factors could conceivably affect track size, because reptile growth is incremental over an animal's life span, because V. niloticus can reach large sizes, and because the reported size of extant reptiles may not be the same as the size attained in the Pleistocene: Klein31 reported that mammalian carnivore size varied significantly during the Pleistocene, being larger during glacial phases, and Tyrberg32 reported that at a global level Late Pleistocene bird species were often larger than their extant descendants. The same phenomenon might apply to reptiles. Nonetheless, 257 cm is greater than the 220 cm upper limit of the V. niloticus size range reported by Keates33. The digit impressions of the large partial track on Surface B are longer than those measured at Surface A. While this may seem to suggest an even larger trackmaker, differences in substrate can also account for such variations. V. albigularis is smaller than V. niloticus, and is even less likely to have attained such a large size. The expert trackers whom we consulted concluded that the large tracks evident on Surface A were consistent with those of C. niloticus. Based on track dimensions, digit shapes, and claw impressions, we conclude that the large tracks on Surface A are consistent with those of the extant C. niloticus or a similar large reptile.
We are not aware of previous reports of fossil reptile swim traces from Africa, although hippopotamus swim traces have been reported.34 We are not aware of reports of fossil non-crocodylian reptile swim traces. However, crocodylian swim traces feature in the global ichnological record.35,36 There is a range of swim trace morphologies, depending on depth of water (and the size and leg length of the swimmer), with tracks made in shallow water at one end of this spectrum (virtually indistinguishable from dryland tracks), and faint, slightly curvilinear impressions made in deep water at the other end. Traces made in relatively deep water are described as 'raking traces', forming parallel or sub-parallel scratch marks.37 Clearly a reptile that is exclusively swimming, and not touching the bottom, leaves no traces whatsoever. Examination of swim traces of extant C. niloticus and V. niloticus would be useful to further document the nature of this spectrum. The flippers of large adult turtles tend to strike the substrate more laterally, and would therefore leave impressions unlike those seen in Figure 6; hence such species can probably be excluded. We interpret the features at Surface C and Surface D as including probable reptile swim traces, while acknowledging that some of the features on Surface C could conceivably be tail drag impressions. Some of the features on Surface C appear consistent with 'raking traces', and therefore appear to have been made in a deeper aqueous environment than those on Surface D, where shorter groove features are contiguous with well-defined digit impressions.
Despite the many smaller tracks and probable swim traces we observed, determining trackmaker identity with confidence proved challenging. Two exceptions were apparent. The partial track shown in Figure 8a and 8b resembles that of a juvenile C. niloticus. The track shown in Figure 4f exhibits a substantial, consistent distance between the anterior ends of the digits and the claw impressions, which led us to conclude that the trackmaker may have been a lizard species similar to the two extant Varanus species.
The fossil and archaeological record
Pickford38 described crocodile remains from Early and Middle Miocene deposits in Namibia. Hendey39 described what is possibly the earliest known occurrence of C. niloticus from Middle Miocene deposits at Arrisdrift, on the Namibian side of the Orange River. Feely40 provided a review of known southern occurrences of C. niloticus, and its current, historical and prehistorical southern range limits. He noted a skull specimen of an unknown saurian found by A.G. Bain in the Eastern Cape at a place Bain called 'Crocodile Hill'. However, the site and geological context are unknown. A large, partially fossilised skull of C. niloticus, of uncertain age, was recovered near the Swartkops estuary east of Port Elizabeth.40
Closer to our study area are two further possible records.40 One is a report of a crocodile tooth found in a shelter at Lottering River (just within the Eastern Cape Province), presumed to have been transported there by a Later Stone Age inhabitant. The second is a report of a crocodile skull from Matjes River, east of Plettenberg Bay, also presumed to have been transported by a Later Stone Age inhabitant. The archaeological context is unknown in both cases.
There is an absence of reports of large reptiles from archaeological sites or contemporary faunal accumulations on the Cape south coast; Richard Klein (2019, written communication, February 15) indicates that in his extensive inventory of the region there are no coastal records of crocodiles or monitor lizards. Literature on Pleistocene reptiles from the Cape south coast from archaeological sites relates to small reptiles (tortoises).41,42
Historical records, folklore and current range limits
West of the tracksites we describe, the Kaaimans River flows into the Indian Ocean. Leggatt43 has researched the history of this name, which could signify 'cayman'. He notes that explorers' accounts from the 18th century use the name with varying spelling, that Le Vaillant in 1782 associated it with crocodiles, and that Lichtenstein in 1803 thought it referred to the water monitor.43 However, these associations were presented without evidence. Leggatt43 provides an alternative derivation, from a Khoisan belief in a giant watersnake, 'Keiman', that inhabited river pools. Feely40 reaches a similar conclusion, while also noting that the name might derive from a smaller lizard.
There are no known historical records of large reptiles from the Cape south coast, nor from adjoining areas. This absence of evidence includes accounts of the coastal regions of the Western Cape and Eastern Cape by survivors of Portuguese shipwrecks between Plettenberg Bay and Pondoland in the 16th and 17th centuries.40
Due to its ectothermic biology, C. niloticus, like most other crocodylians, is limited to tropical and subtropical areas with daily maximum ambient temperatures of 25-35 °C.44 The southward cooling of habitats forms the most significant determinant of its range limits. All recorded localities for this species towards the southern limits of its range are at or near coastal estuaries or wetlands. The closest C. niloticus population to the tracksites described here is at the Dwesa-Cwebe Nature Reserve in the Eastern Cape Province.40,45 Six juvenile C. niloticus were introduced from Zululand to this area in 1977,45 and two successful breeding events have been documented (Venter J 2014, written communication).
Less information is available on the historical or current distribution of V. niloticus, although similar considerations can be assumed regarding the southward cooling of temperature. Lubke and De Moor46 report that it extends southwest as far as Kouga River, Eastern Cape. Keates33 reports the southern range limit at Seekoei River, Eastern Cape. Both localities are more than 200 km east of the tracksites we describe.
Lithics associated with fossil tracks
Although observations of the artifacts on Surface B are necessarily preliminary (Figure 5), their characteristics suggest that they date from the MSA.47-50 We are not aware of prior reports of lithics embedded in Pleistocene palaeosurfaces from the Cape south coast, and certainly none that occur in association with fossil tracks. Roberts et al.1 reported 'six non-diagnostic stone artefacts' from a palaeosol between aeolianite layers east of Still Bay. Graham Avery (2019, written communication, February 10) reports occurrences of artifacts in association with aeolianite outcrops from False Bay (at the Holocene / Late Pleistocene erosional surface in the Olympic Sand Mine). From the West Coast, Cruz-Uribe et al.51 reported Late Acheulean artifacts in palaeosols between calcareous deposits dating to as much as 160 ka at Duinefontain, and Avery et al.52 reported artifacts and shell middens in an eroded overhang dated to MIS 5 at Ysterfontein. To the east, Kevin Cole (2019, written communication, February 5) reports blades and scrapers embedded in an aeolianite surface, near the Nahoon Point hominin tracks, and Graham Avery (2019, written communication, February 10) reports having found a mSa convergent flake in this same area. Pargeter et al.53 described lithics in Middle Pleistocene dune surfaces from the Pondoland coast. The significance of such occurrences may be greater if they are found on the same surfaces as fossil tracks.
We found no evidence to suggest that any tracks on Surface B were of hominin origin, and the lithics were embedded in the layer of sediment that appeared to have washed over the track-bearing surface. Nonetheless, a spatial and temporal association in this environment between humans and large reptiles can be inferred, or at least mutual use of habitat, something which has not previously been documented. The MSA date for the lithics is consistent with the range of OSL dates obtained from nearby deposits.
The loss of this site to further study as a result of a landslide is regrettable. However, this is consistent with the ephemeral nature of many tracksites on the Cape south coast. It underlines the importance of early detection and documentation of newly exposed tracksites.
Palaeoenvironmental considerations
For C. niloticus embryos to hatch successfully, incubation (i.e. sand) temperatures must be sufficiently warm (28-35 °C) for egg incubation.54 Measurement of sand temperatures close to the sites we describe would establish whether or not egg incubation is currently theoretically possible on the Cape south coast. We suspect that it is not, and that the reptile tracks were registered during a warmer period during the Pleistocene during or close to the time of a sea highstand that allowed reptile species to occupy an extended range. Chronological context11 suggests an age of MIS 5e (the Last Interglacial). As sea levels during MIS 5e in this area were up to 6-8 m higher than at present, a warmer climate capable of supporting large reptiles on the Cape south coast can be inferred.20
Conclusions
The sites we describe represent the first evidence of large reptiles (such as C. niloticus and V. niloticus) on the Cape south coast of South Africa, and probably the first recorded reptile swim traces in Africa. They join the substantial record of avian and mammal Pleistocene tracksites along this coastline, and complement the extensive record of body fossils. Their importance includes palaeoenvironmental implications, as they were probably registered during a sea highstand characterised by warm temperatures - consistent with the ectothermic biological requirements of large reptiles such as C. niloticus and probably V. niloticus. The finding of two MSA artifacts on one of the track-bearing surfaces represents a previously undocumented association of Pleistocene hominin populations with large reptiles. Further studies could include recovery and more detailed analysis of the artifacts, measurement of sand temperatures close to the tracksites, and documentation of the swim traces of extant large African reptiles.
Acknowledgements
We acknowledge the support of Graham Avery, Andre Brink, Emily Brink, Kevin Cole, Jessica Hayes and staff of Garden Route National Park, Linda Helm, Richard Klein, Martin Lockley, Liezl Van Pletzen-Vos and Jan Venter. We thank expert trackers Ian Thomas and Alan Yeowart for their valuable opinions.
Authors' contributions
C.W.H.: Lead author, corresponding author, conceptualisation, data collection, data analysis, writing, project leadership, track analysis. H.C.C.: Conceptualisation, sample analysis, site visit, data analysis, contribution on geological context, field stratigraphy, review of drafts and revisions. X.C.: Conceptualisation, data analysis, site visit, contributions on crocodile records and reptile biology, review of drafts and revisions. C.J.Z.H.: Conceptualisation, data analysis, photogrammetry, review of drafts and revisions. R.R.: Conceptualisation, data analysis, site visit, contributions on MSA lithics, review of drafts and revisions. W.S.: Conceptualisation, data analysis, site visits, contributions on sedimentology, review of drafts and revisions. A.v.d.H.: Conceptualisation, data analysis, site visit, contributions on ichnology and track analysis, review of drafts and revisions.
References
1. Roberts DL, Bateman MD, Murray-Wallace V Carr AS, Holmes PJ. Last interglacial fossil elephant trackways dated by OSL/AAR in coastal aeolianites, Still Bay, South Africa. Palaeogeogr Palaeoclimatol Palaeoecol. 2008;257(3):261-279. http://dx.doi.org/10.1016/j.palaeo.2007.08.005 [ Links ]
2. Helm CW, Anderson RJ, Buckley LG, Cawthra HC, De Vynck JC. Biofilm assists recognition of avian trackways in Late Pleistocene coastal aeolianites, South Africa. Palaeont Afr. 2017;52:78-84. [ Links ]
3. Helm CW, Cawthra HC, Cowling RM, De Vynck JC, Marean CW, McCrea RT, et al. Palaeoecology of giraffe tracks in Late Pleistocene aeolianites on the Cape south coast. S Afr J Sci. 2018;114(1/2), Art. #2017-0266, 8 pages. http://dx.doi.org/10.17159/sajs.2018/20170266 [ Links ]
4. Helm CW, McCrea RT, Cawthra HC, Thesen GHH, Mwankunda JM. Late Pleistocene trace fossils in the Goukamma Nature Reserve, Cape south coast, South Africa. Palaeont Afr. 2018;52:89-101. [ Links ]
5. Helm CW, McCrea RT, Cawthra HC, Cowling RM, Lockley MG, Marean CW, et al. A new Pleistocene hominin tracksite from the Cape south coast, South Africa. Sci Rep. 2018;8, Art. #3772, 13 pages. http://dx.doi.org/10.1038/s41598-018-22059-5 [ Links ]
6. Helm CW, Lockley MG, Cole K, Noakes TD, McCrea RT. Hominin tracks in southern Africa: A review and an approach to identification. Palaeont Afr. 2019;53:81-96. [ Links ]
7. Helm CW, Cawthra HC, De Vynck JC, Lockley MG, McCrea RT, Venter J. The Pleistocene fauna of the Cape south coast revealed through ichnology at two localities. S Afr J Sci. 2019;115(1/2), Art. #5135, 9 pages. https://doi.org/10.17159/sajs.2019/5135 [ Links ]
8. Klein RG. The late quaternary mammalian fauna of Nelson Bay Cave (Cape Province, South Africa): Its implications for megafaunal extinctions and environmental and cultural change. Quat Res. 1972;2:135-142. https://doi.org/10.1016/0033-5894(72)90034-8 [ Links ]
9. Klein RG. The mammalian fauna of the Klasies River Mouth sites, southern Cape Province, South Africa. S Afr Archaeol Bull. 1976;31:75-98. https://doi.org/10.2307/3887730 [ Links ]
10. Klein RG. Palaeoenvironmental implications of Quaternary large mammals in the Fynbos Region. In: Deacon HJ, Hendey QB, Lambrechts JJN, editors. Fynbos palaeoecology: A preliminary synthesis. Pretoria: Cooperative Scientific Programmes; 1983. p. 116-138. [ Links ]
11. Bateman MD, Carr AS, Dunajko AC, Holmes PJ, Roberts DL, Mclaren SJ, et al. The evolution of coastal barrier systems: A case study of the Middle-Late Pleistocene Wilderness barriers, South Africa. Quat Sci Rev. 2011;30:63-81. https://doi.org/10.1016/j.quascirev.2010.10.003 [ Links ]
12. Roberts DL, Karkanas P Jacobs Z, Marean CW, Roberts RG. Melting ice sheets 400,000 yr ago raised sea level by 13 m: Past analogue for future trends. Earth Planet Sci Lett. 2012;357-358:226-237. https://doi.org/10.1016/j.epsl.2012.09.006 [ Links ]
13. Cawthra HC, Jacobs Z, Compton JS, Fisher EC, Karkanas P Marean CW. Depositional and sea-level history from MIS 6 (Termination II) to MIS 3 on the southern continental shelf of South Africa. Quat Sci Rev. 2018;181:156-172. https://doi.org/10.1016/j.quascirev.2017.12.002 [ Links ]
14. Carr AS, Bateman MD, Cawthra HC, Sealy J. First evidence for onshore marine isotope stage 3 aeolianite formation on the southern Cape coastline of South Africa. Mar Geol. 2019;407:1-15. https://doi.org/10.1016/j.margeo.2018.10.003 [ Links ]
15. Malan JA. Lithostratigraphy of the Waenhuiskrans Formation (Bredasdorp Group). South African Committee for Stratigraphy Lithostratigraphic Series 8. Pretoria: Department of Mineral and Energy Affairs; 1989. [ Links ]
16. Malan JA. Lithostratigraphy of the Klein Brak Formation (Bredasdorp Group). South African Committee for Stratigraphy Lithostratigraphic Series 13. Pretoria: Department of Mineral and Energy Affairs; 1991. [ Links ]
17. Illenberger WK. The geomorphic evolution of the Wilderness dune cordons, South Africa. Quat Int. 1996;33:11-20. https://doi.org/10.1016/1040-6182(95)00099-2 [ Links ]
18. Bateman MD, Holmes PJ, Carr AS, Horton BP Jaiswal MK. Aeolianite and barrier dune construction spanning the last two glacial-interglacial cycles from the southern Cape coast, South Africa. Quat Sci Rev. 2004;23:1681-1698. https://doi.org/10.1016/j.quascirev.2004.02.001 [ Links ]
19. Fisher EC, Bar-Matthews M, Jerardino A, Marean CW. Middle and Late Pleistocene paleoscape modeling along the southern coast of South Africa. Quat Sci Rev. 2010;29:1382-1398. http://dx.doi.org/10.1016/jquascirev.2010.01.015 [ Links ]
20. Carr AS, Bateman MD, Roberts DL, Murray-Wallace CV Jacobs Z, Holmes PJ. The last interglacial sea-level high stand on the southern Cape coastline of South Africa. Quat. Res. 2010;73:351-363. https://doi.org/10.1016/j.yqres.2009.08.006 [ Links ]
21. Cawthra HC, Cowling RM, Andò S, Marean CW. Geological and soil maps of the Palaeo-Agulhas Plain for the Last Glacial Maximum. Quat Sci Res. In press 2019. https://doi.org/10.1016/j.quascirev.2019.07.040 [ Links ]
22. Stuart C, Stuart T. A field guide to the tracks and signs of southern and East African wildlife. Cape Town: Struik Nature; 2000. [ Links ]
23. Matthews NA, Noble TA, Breithaupt BH. Close-range photogrammetry for 3-D ichnology: The basics of photogrammetric ichnology. In: Falkingham PL, Marty D, Richter A, editors. Dinosaur tracks: The next steps. Bloomington, IN: Indiana University Press; 2016. p. 28-55. [ Links ]
24. Liebenberg L. A photographic guide to tracks and tracking in southern Africa. Cape Town: Struik Publishers; 2000. [ Links ]
25. Van den Heever A, Mhlongo R, Benadie K. Tracker manual - A practical guide to animal tracking in southern Africa. Cape Town: Struik Nature; 2017. [ Links ]
26. Branch B. Field guide to the snakes and other reptiles of southern Africa. Cape Town: Struik; 1994. [ Links ]
27. Marean CW, Cawthra HC, Cowling RM, Esler KJ, Fisher E, Milewski A, et al. Stone Age people in a changing South African Greater Cape Floristic Region. In: Allsopp N, Colville JF, Verboom GA, editors. Fynbos: Ecology, evolution, and conservation of a megadiverse region. Oxford: Oxford University Press; 2014. p. 164-199. http://dx.doi.org/10.1093/acprof:oso/9780199679584.001.0001 [ Links ]
28. Cawthra HC. The marine geology of Mossel Bay [PhD thesis]. Cape Town: University of Cape Town; 2014. [ Links ]
29. Nichols G. Sedimentology and stratigraphy. Chichester; John Wiley and Sons Ltd.; 2009. [ Links ]
30. Thomson WR. A field guide for estimating the overall length of the crocodile. S Afr J Wildl Res. 1972;2(1):27. [ Links ]
31. Klein RG. Carnivore size and Quaternary climatic change in southern Africa. Quat Res. 1986;26:153-170. https://doi.org/10.1016/0033-5894(86)90089-X [ Links ]
32. Tyrberg T. The Late Pleistocene continental avian extinction - An evaluation of the fossil evidence. Oryctos. 2007;7:249-269. [ Links ]
33. Keates C. The monitor lizards of southern Africa [webpage on the Internet]. c2016 [cited 2019 Nov 08]. Available from: https://nextgenherpetologist.co.za/2016/12/23/the-monitor-lizards-of-southern-africa/ [ Links ]
34. Bennett MR, Morse SA, Falkingham PL. Tracks made by swimming hippopotami: An example from Koobi Fora (Turkana Basin, Kenya). Palaeogeogr Palaeoclimatol Palaeoecol. 2014;409:9-23. http://dx.doi.org/10.1016/j.palaeo.2014.04.021 [ Links ]
35. Lockley MG, Lucas SG, Milan J, Harris JD, Avanzini M, Foster JR, et al. The fossil record of crocodylian tracks and traces: An overview. In: Milan J, Lucas SG, Lockley MG, Spielmann JA, editors. Crocodyle tracks and traces. New Mexico Museum of Natural History and Science Bulletin 51. Albequerque, NM: New Mexico Museum of Natural History and Science; 2010. p. 1-15. [ Links ]
36. Lockley MG, Lucas SG. Fossil footprints of western North America. New Mexico Museum of Natural History and Science Bulletin 62. Albequerque, NM: New Mexico Museum of Natural History and Science; 2014. [ Links ]
37. Romano M, Whyte MA. Crocodilian and other non-dinosaurian tracks and trackways from the Ravenscar Group (Middle Jurassic) of the Cleveland Basin, Yorkshire, UK. In: Milan J, Lucas SG, Lockley MG, Spielmann JA, editors. Crocodyle tracks and traces. New Mexico Museum of Natural History and Science Bulletin 51. Albequerque, NM: New Mexico Museum of Natural History and Science; 2010. p. 69-81. [ Links ]
38. Pickford M. A new species of crocodile from Early and Middle Miocene deposits in the lower Orange River Valley, Namibia and the origins of the Nile crocodile (Crocodylus niloticus). In: Pickford M, Senut B, editors. Memoir of the Geological Survey of Namibia: Geology and Palaeobiology of the Central and Southern Namib: Palaeontology of the Orange River Valley, Namibia. Volume 19. Windhoek: Geological Survey of Namibia; 2003. p. 51-65. [ Links ]
39. Hendey QB. Preliminary report on the Miocene vertebrates from Arrisdrift, South West Africa. Ann S Afr Mus. 1978;76:1-11. [ Links ]
40. Feely JM. On the southeastern range limits of the Nile crocodile: A review of its past and present occurrences in the Eastern Cape and Western Cape, South Africa. S Afr J Wildl Res. 2010;40(2):169-175. https://doi.org/10.3957/056.040.0209 [ Links ]
41. Reynard JP, Henshilwood CS. Subsistence strategies during the Late Pleistocene in the southern Cape of South Africa: Comparing the Still Bay of Blombos Cave with the Howiesons Poort of Klipdrift Shelter. J Hum Evol. 2017;108:110-130. https://doi.org/10.1016/j.jhevol.2017.04.003 [ Links ]
42. Thompson JC, Henshilwood CS. Tortoise taphonomy and tortoise butchery patterns at Blombos Cave, South Africa. J Archaeol Sci. 2014;41:214-229. https://doi.org/10.1016/j.jas.2013.08.017 [ Links ]
43. Leggatt H. Wilderness names [document on the Internet]. c2016 [cited 2019 Nov 08]. Available from: http://wrra.co.za/images/stories/PDF/HugoLeggatt/WildernessX20Names.pdf [ Links ]
44. Grigg G, Kirshner D. Biology and evolution of crocodylians. New York: Cornell University Press; 2015. [ Links ]
45. Pooley AC. Crocodile research in Maputaland. In: Bruton MN, Cooper KH, editors. The ecology of Maputaland. Cape Town: Cape & Transvaal Printers (Pty) Ltd; 1980. p. 293-299. [ Links ]
46. Lubke R, De Moor I. Guide to the eastern and southern Cape coasts. Cape Town: UCT Press; 1998. [ Links ]
47. Wurz S. Middle Stone Age tools from Klasies River main site, conventions and symbolic cognition. In: Nowell A, Davidson I, editors. The cutting edge: Stone tools and the evolution of cognition. Boulder, CO: Colorado University Press; 2010. p. 135-158. [ Links ]
48. Wurz S. New investigations at Klasies River Main site. The Digging Stick. 2016;33(3):7-10. [ Links ]
49. Brown KS, Marean CW, Herries AIR, Jacobs Z, Tribolo C, Braun D, et al. Fire as an engineering tool of early modern humans. Science. 2009;325:859-862. http://dx.doi.org/10.1126/science.1175028 [ Links ]
50. Lombard M. Evidence for hunting and hafting at Sibudu Cave during the Middle Stone Age. The Digging Stick. 2002;19(3):1-3. [ Links ]
51. Cruz-Uribe K, Klein RG, Avery G, Avery DM, Halkett D, Hart T, et al. Excavation of buried Late Acheulean (Mid-Quaternary) land surfaces at Duinefontein 2, Western Cape Province, South Africa. J Archaeol Sci. 2003;30(5):559-575. https://doi.org/10.1016/S0305-4403(02)00202-9 [ Links ]
52. AveryG, Halkett D, Orton JDJ, Steele TE, Tusenius M, Klein RG. The Ysterfontein 1 Middle Stone Age rock shelter and the evolution of coastal foraging. Goodwin Ser. 2007;10:66-89. https://www.jstor.org/stable/40650020 [ Links ]
53. Pargeter J, Saktura R, Blackwood A, Cawthra H, Esteban I, Fisher E, et al. The Msikaba Red Sand Dunes: Middle Pleistocene lithic technological variability in Pondoland, South Africa. Paper presented at: The 84th Annual Meeting of the Society for American Archaeology; 2019 April 10-14; Albequerque, New Mexico, USA. [ Links ]
54. Hutton JM. Incubation temperatures, sex ratios and sex determination in a population of Nile crocodiles (Crocodylus niloticus). J Zool. 1987;211:143-155. https://doi.org/10.1111/j.1469-7998.1987.tb07458.x [ Links ]
 Correspondence:
Correspondence:
Charles Helm
helm.c.w@gmail.com
Received: 19 July 2019
Revised: 08 Nov. 2019
Accepted: 08 Nov. 2019
Published: 26 Mar. 2020
Editor: Maryna Steyn
Funding: None

{kind=link}